Класс млекопитающие

Появление настоящих млекопитающих произошло в мезозойскую эру. Десятки миллионов лет млекопитающие ютились на задворках мира динозавров, но, в конечном счете, эволюция расставила все на свои места: изменения климата привели к гибели динозавров. Здесь то и восторжествовали теплокровные млекопитающие, которые к настоящему времени заселили весь мир.
Ароморфозы млекопитающих
Замечу, что ароморфозы млекопитающих возникли независимо от птиц: это две разные эволюционные ветви. Млекопитающие произошли не от птиц, а от древних пресмыкающихся. Поэтому не удивляйтесь, если некоторые ароморфозы окажутся уже вам знакомы.
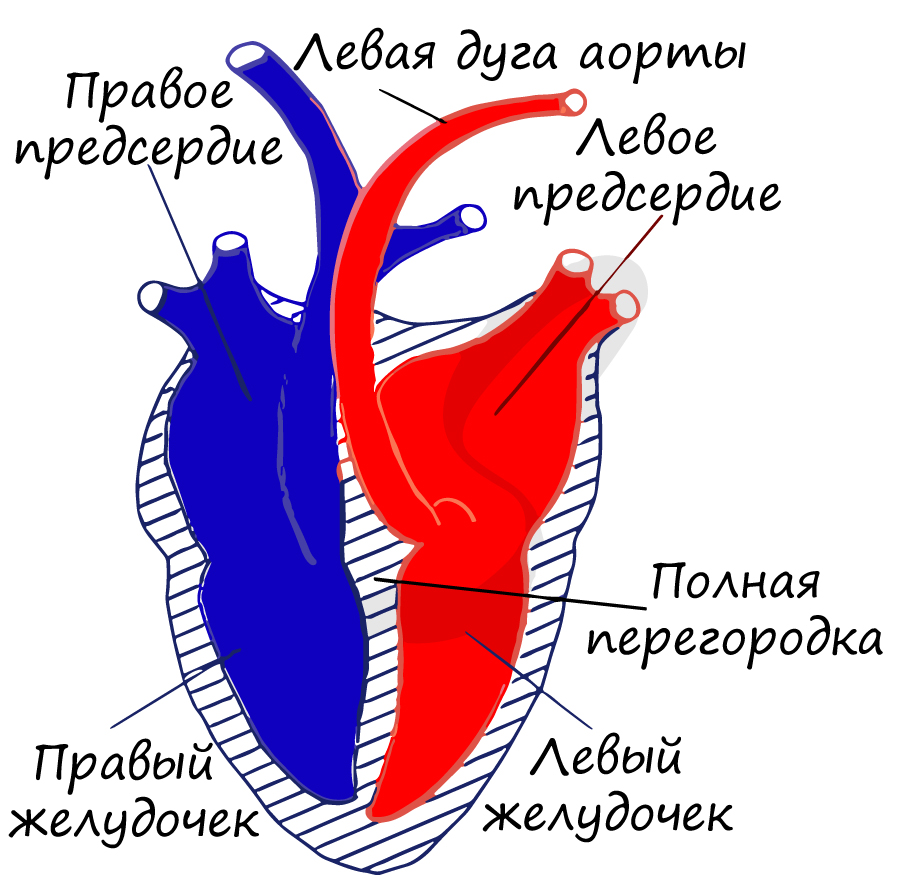
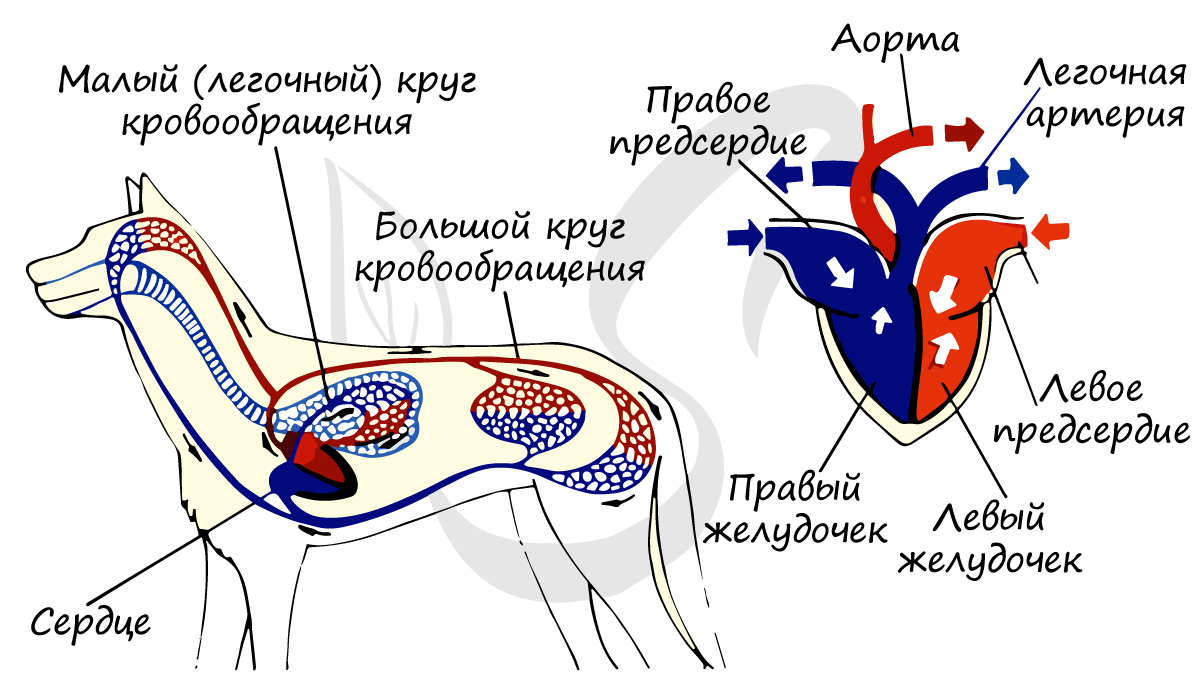
У млекопитающих в сердце возникает полная перегородка: кровь не смешивается, два круга кровообращения полностью отделены друг от друга. Это значительно повышает уровень обмена веществ: температура тела млекопитающих более не зависит от температуры окружающей среды, они приобретают теплокровность (гомотермию).
Развитие коры больших полушарий (КБП), центра высшей нервной деятельности, достигает пика. Такое сильное развитие КБП обеспечивает возникновение сложного поведения, облегчает формирование условных рефлексов, способствуя повышению выживаемости вида.
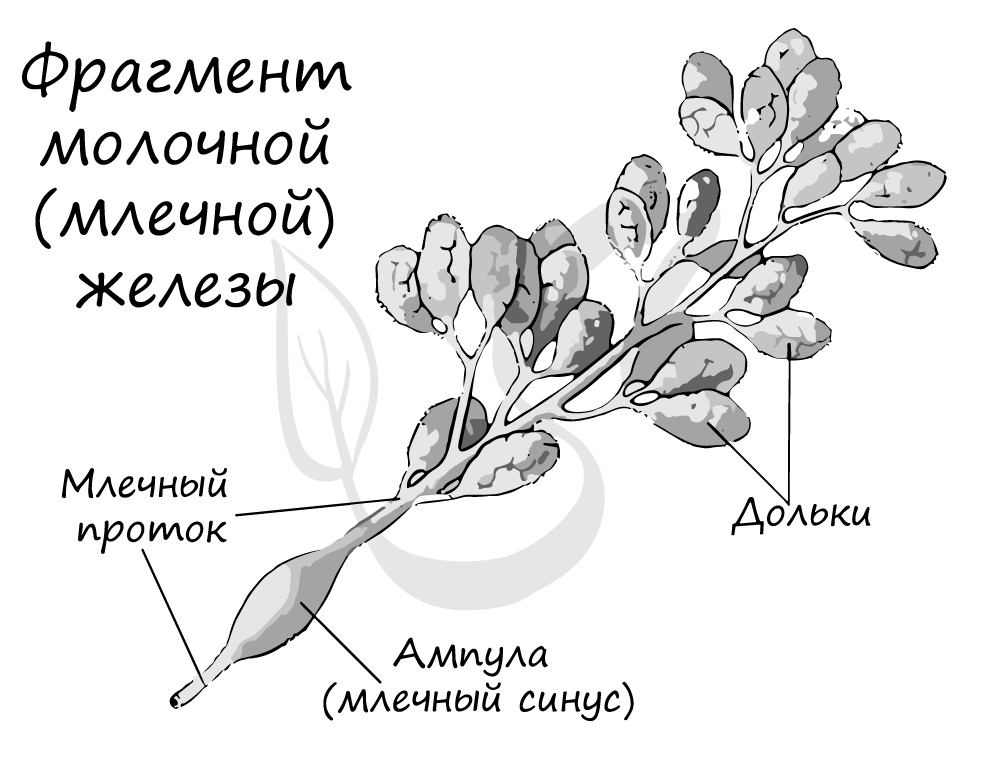
Для млекопитающих характерна забота о потомстве, проявляющаяся и в уже упомянутом вскармливании молоком, и в том, что детеныши часто живут с матерью несколько лет. Еще во время беременности самка ищет укромное место для рождения потомства, заранее заготавливает корм. Такая забота выглядит удивительно трогательно в диком животном мире.

Волосяной покров помогает более эффективно сохранять тепло, обеспечивая термоизоляцию организму животного. Часть млекопитающих утратила волосяной покров, в связи со вторичноводным образом жизни: они избрали своей средой обитания моря, океаны, где волосяной покров явно был бы лишним и создавал ненужное трение о воду.

По выполняемой функции зубы делятся на резцы, клыки, предкоренные и коренные зубы. У некоторых групп животных, к примеру, у грызунов резцы приобретают способность к неограниченному росту. Часто хищники имеют хорошо развитые клыки: тигры, львы.

Классификация млекопитающих
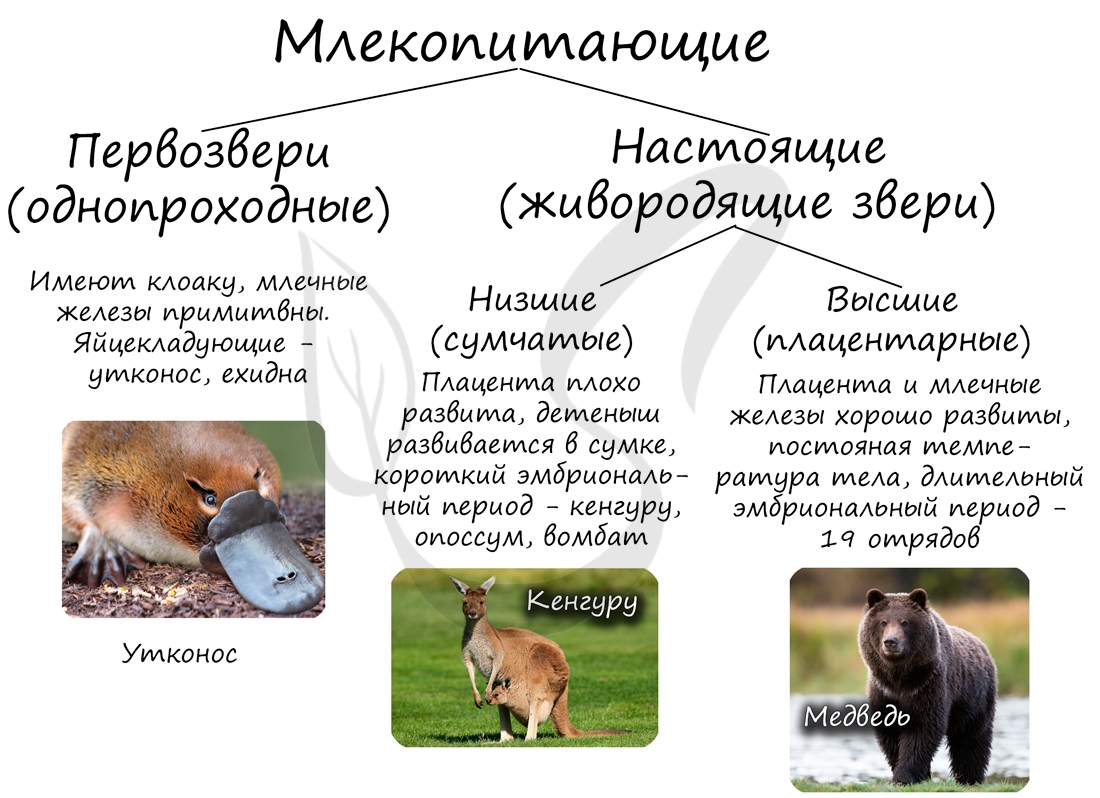
Млекопитающие делятся на подкласс первозверей и противопоставленных им зверей, к которым относятся сумчатые и плацентарные.

Домашняя собака
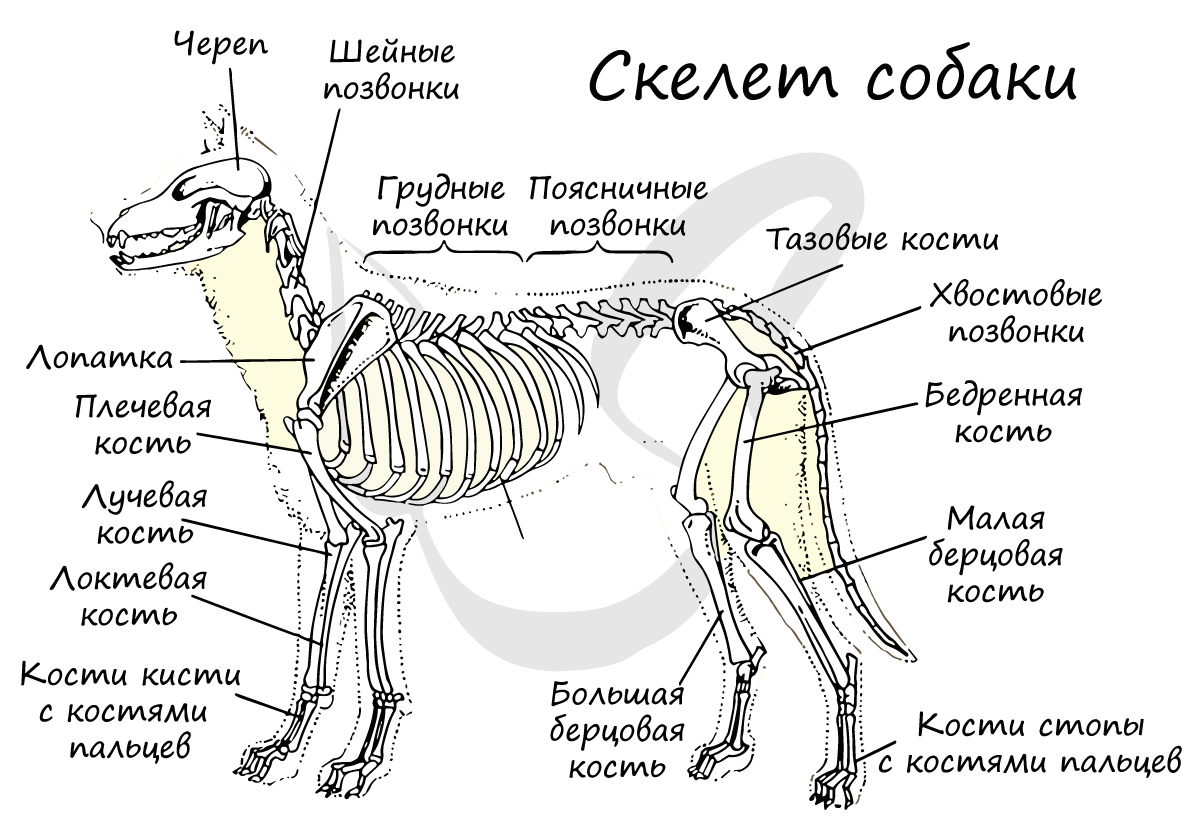
Тело подразделяется на голову, туловище и хвост. Имеются парные конечности: передние и задние, расположенные под телом животного, что является отличием от рептилий, у которых конечности расположены по бокам тела.
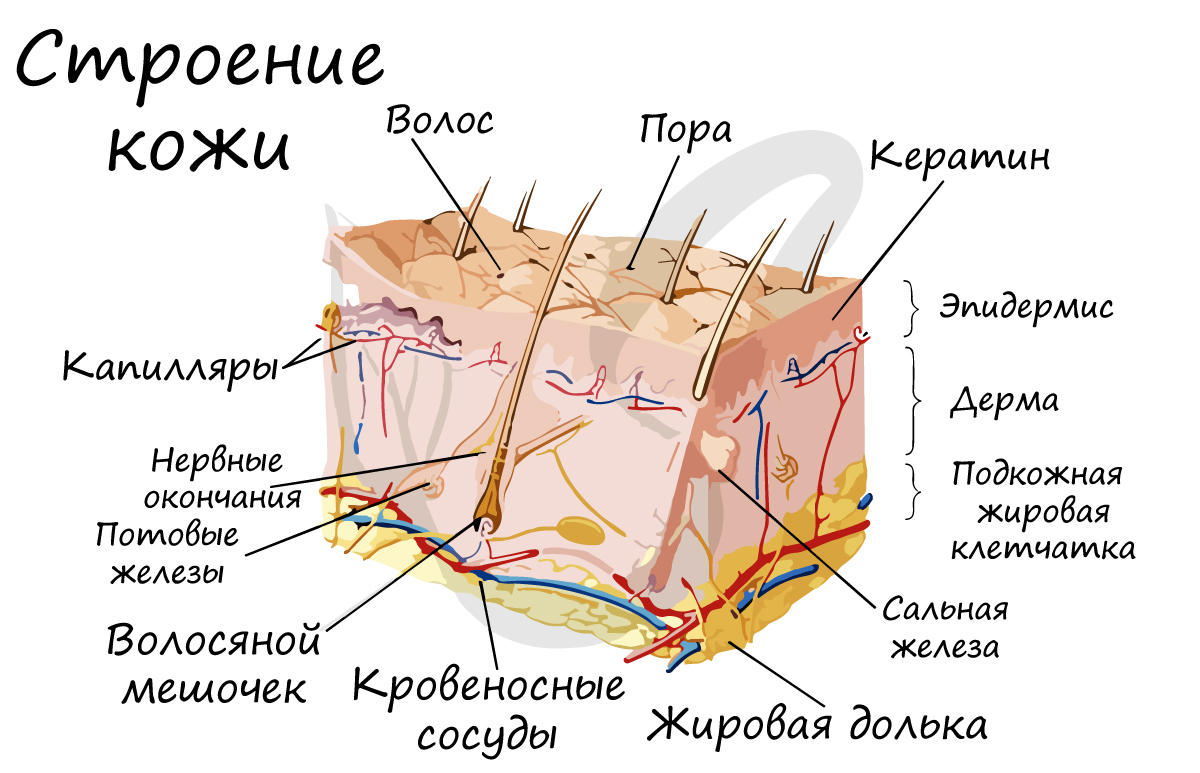
Две пары неубирающихся когтей (производных эпидермиса) расположены на конечностях. Кожа собаки эластичная и прочная, тело покрыто волосяным покровом, состоящим из ости и подшерстка.

В толще кожи проходит большое количеств кровеносных капилляров, которые участвуют в терморегуляции: могут сужаться (во время холода), либо расширяться (во время жары). Таким образом, кожа является депо крови и принимает участие в терморегуляции.
В коже находятся сальные железы, протоки которых открываются в волосяные сумки. Сальные железы выделяют жироподобный секрет, покрывающий кожу защитным слоем. Потовые железы имеют вид трубочек, свернутых в клубок, протоки которых открываются на поверхность кожи. При испарении секрета потовых желез поверхность кожи охлаждается, таким образом, потовые железы участвуют в терморегуляции.
Позвоночник состоит из позвонков с плоской поверхностью, между которыми лежат хрящевые диски, помогающие равномерно распределить нагрузку на позвонки. Позвоночник включает 5 отделов: шейный (7 позвонков), грудной (12-15), поясничный (2-9), крестцовый (1-9) и хвостовой (3-49).
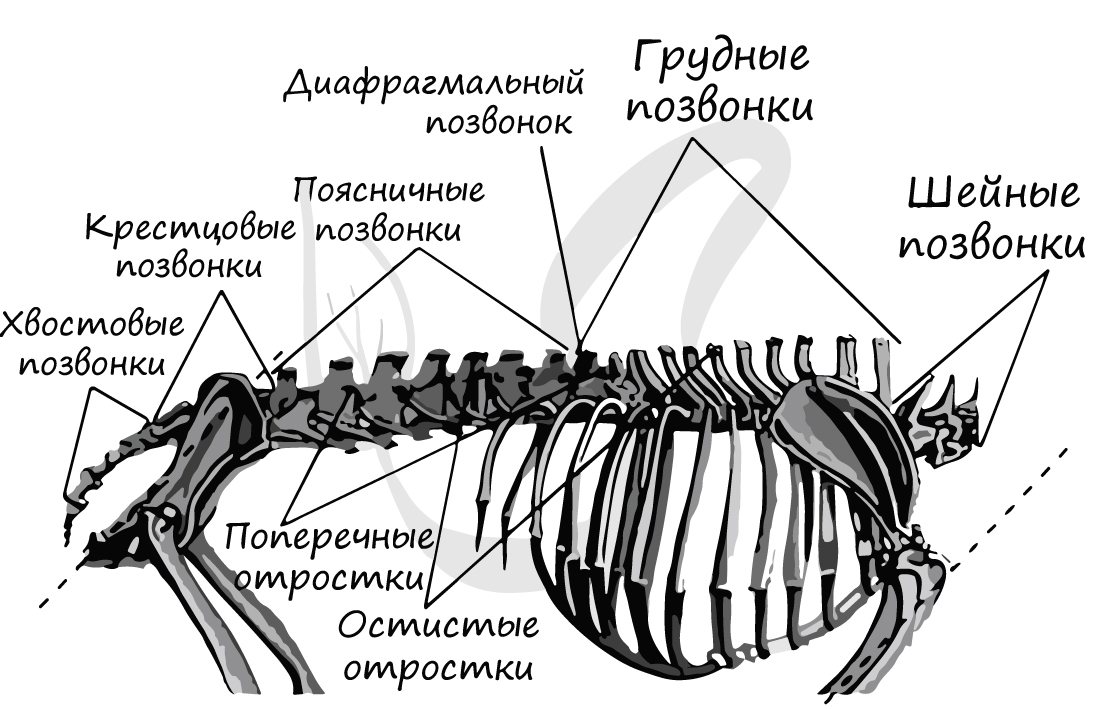
Пояс передних конечностей (плечевой) представлен ключицами и лопатками, коракоид подвергается редукции (исчезновению). Пояс задних конечностей (тазовый) состоит из тазовых костей: седалищной, подвздошной и крестцовой.
Свободная передняя конечность представлена плечевой костью (плечо), локтевой и лучевой костями (предплечье), запястьем, пястьем и фалангами пальцев (кисть). Скелет свободной задней конечности включает в себя бедренную кость (бедро), малую и большую берцовые кости (голень), предплюсну, плюсну и фаланги пальцев (стопу).
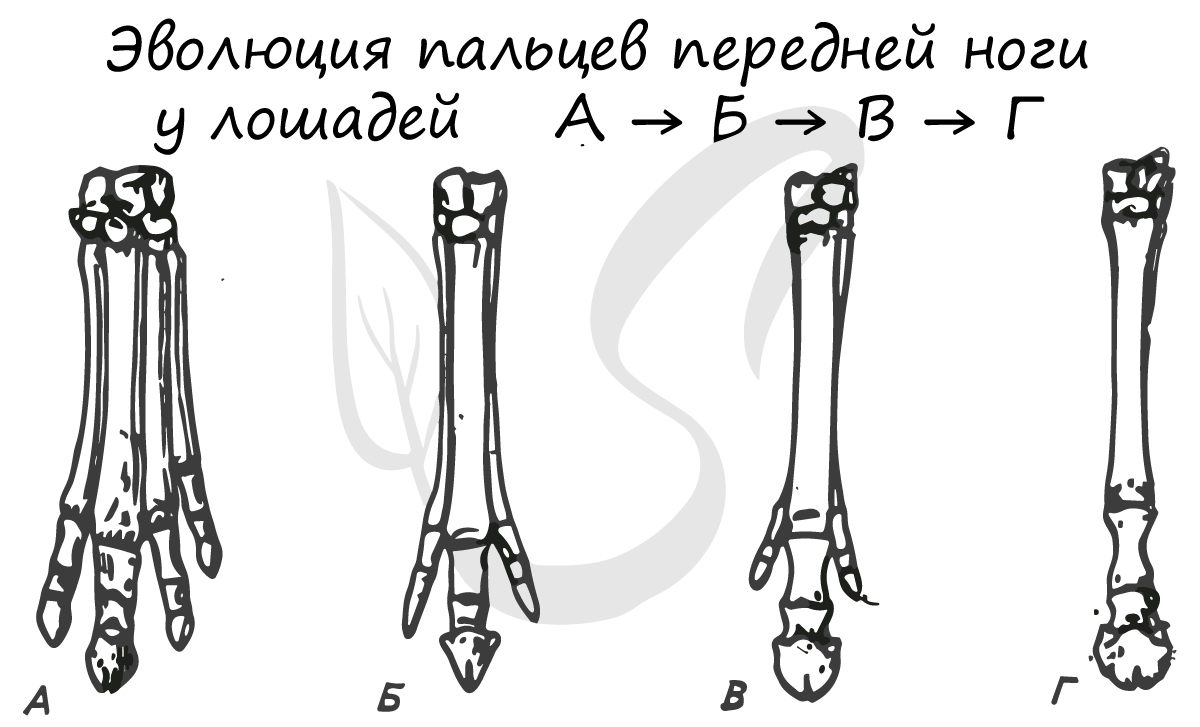
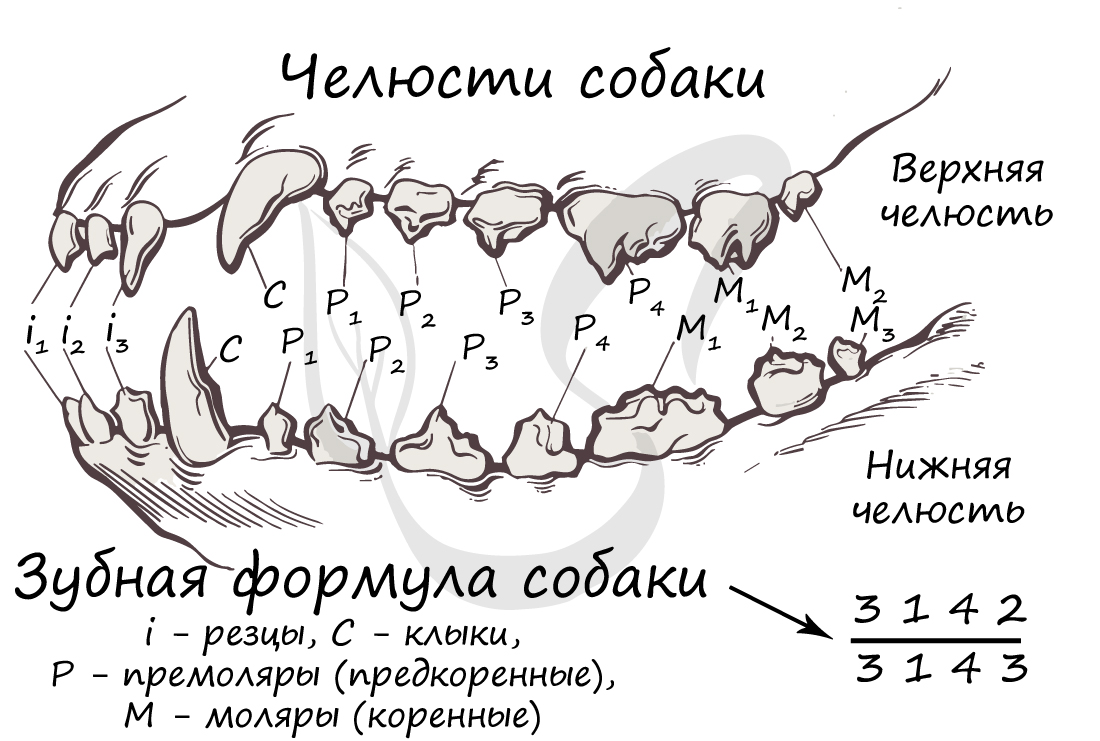
Зубная формула может быть составлена для любого вида зверей, в зависимости от образа жизни она будет отличаться. Верхняя часть дроби отражает строение верхней челюсти, нижняя, соответственно, нижней. Числа обозначают количество зубов в последовательности: резцы, клыки, премоляры (предкоренные) и моляры (коренные).
В ротовую полость открываются слюнные железы. Пищеварительная система в целом типичного строения, в тонкую кишку открываются протоки печени и желчного пузыря, поджелудочной железы.
Особо необходимо отметить желудок жвачных животных. Из-за больших объемов растительной пищи, потребляемой ими, желудок имеет сложное строение. Он состоит из 4 отделов: сычуг, книжка, сетка и рубец.

Сначала пища попадает в рубец, где под действием микроорганизмов происходит расщепление целлюлозы: такая картина напоминает бродильный чан. Далее пища отрыгивается и вторично заглатывается, на этот раз, попадая сначала в сетку, затем в книжку и, наконец, в сычуг, который продолжается в тонкий кишечник.
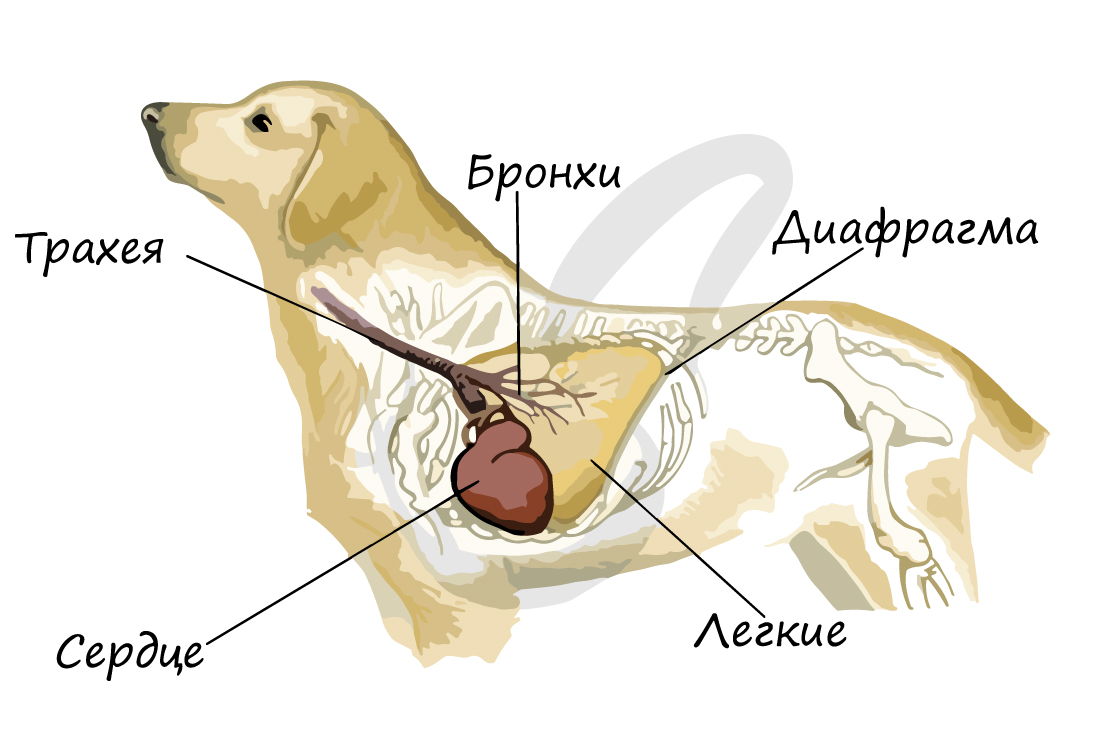
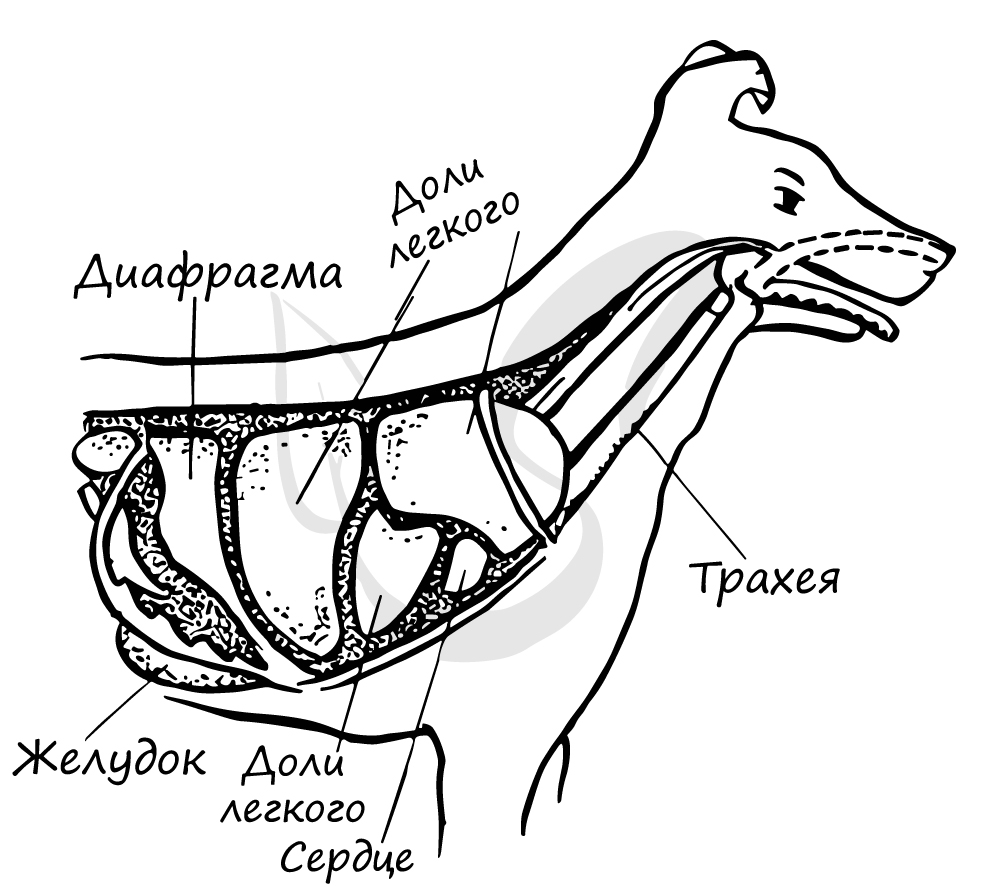
Дыхательная система участвует также в терморегуляции: по мере продвижения через воздухоносные пути воздух нагревается. При учащении дыхания интенсивнее идет теплоотдача.
Животные со слабо развитыми потовыми железами охлаждают свой организм, испаряя воду с поверхности языка. Так у собак в жаркую погоду частота дыхательных движений может достигать 30 в минуту.
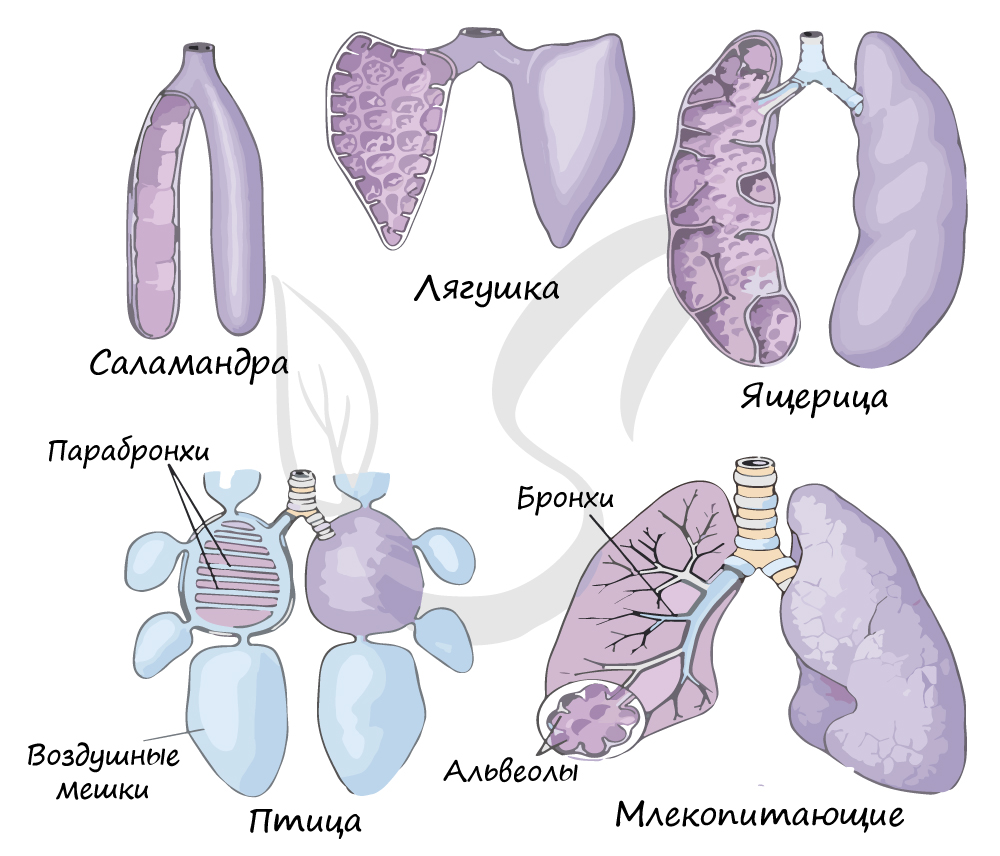
Теперь, когда вами изучены классы хордовых, вы можете познать эволюцию в сравнении. На схеме ниже вы увидите, как постепенно усложнялось строение легких, увеличивалась дыхательная поверхность.

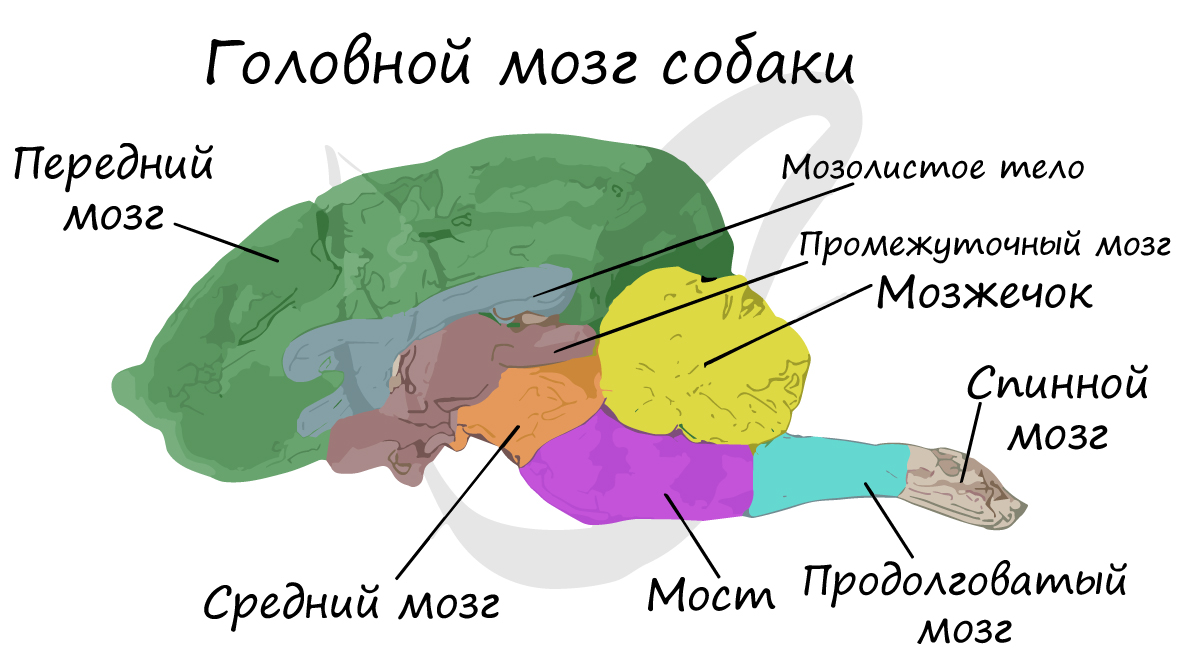
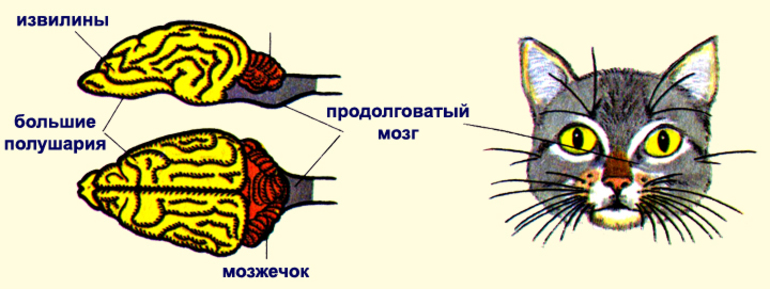
Вам уже известно, что нервная система достигает исключительно высокого уровня развития. Сложное поведение млекопитающих и быстрое легкое формирование у них условных рефлексов осуществляется в первую очередь благодаря развитию коры больших полушарий головного мозга.
Благодаря бороздам, углублениям участков головного мозга, и извилинам, волнистым складкам, поверхность больших полушарий значительно увеличивается. Такой тип мозга называется кортикальный (от лат. cortex — кора). Хорошо развит мозжечок, отвечающий за координацию движений. Средний мозг относительно небольших размеров.
Органы чувств у разных отрядов млекопитающих развиты неодинаково. Органы обоняния находятся в верхней части носовой полости, хорошо развиты у собак: в их головном мозге можно обнаружить две большие обонятельные доли.
Органы зрения представлены глазами. Аккомодация, настройка глаза на наилучшее видение объекта, у млекопитающих достигается только изменением кривизны хрусталика под влиянием сокращений ресничной мышцы.
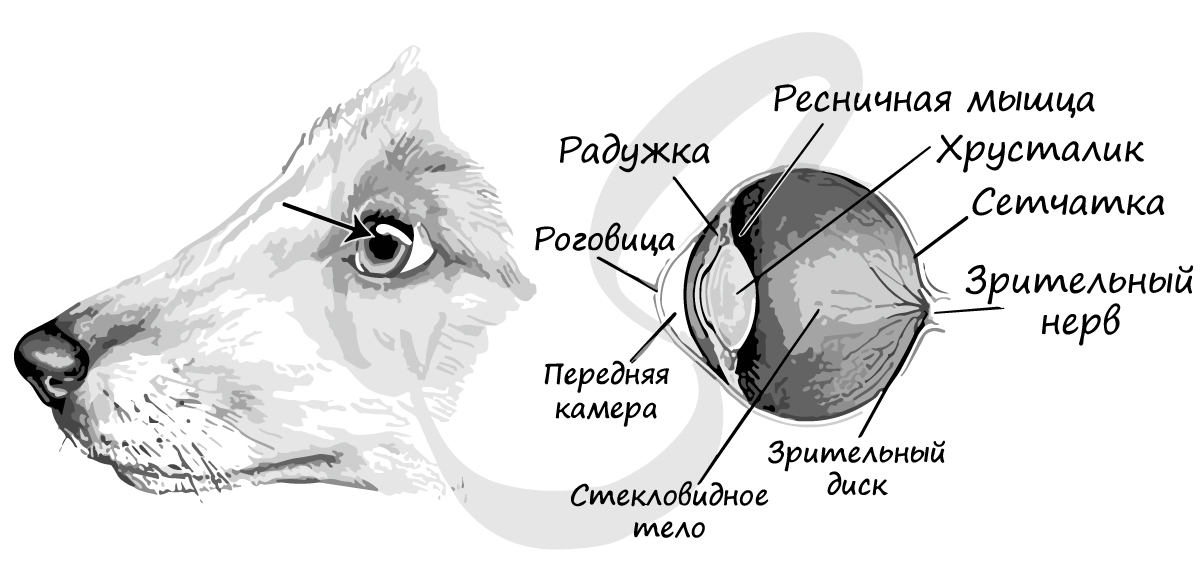
Среднее ухо содержит уже три слуховых косточки: молоточек, наковальню и стремечко.

Вибриссы улавливают воздушные потоки, которые отражаются от объектов окружающей среды. С помощью вибрисс и при участии органов зрения головной мозг строит картину трехмерного мира, благодаря чему животное в полной темноте может успешно обходить препятствия.

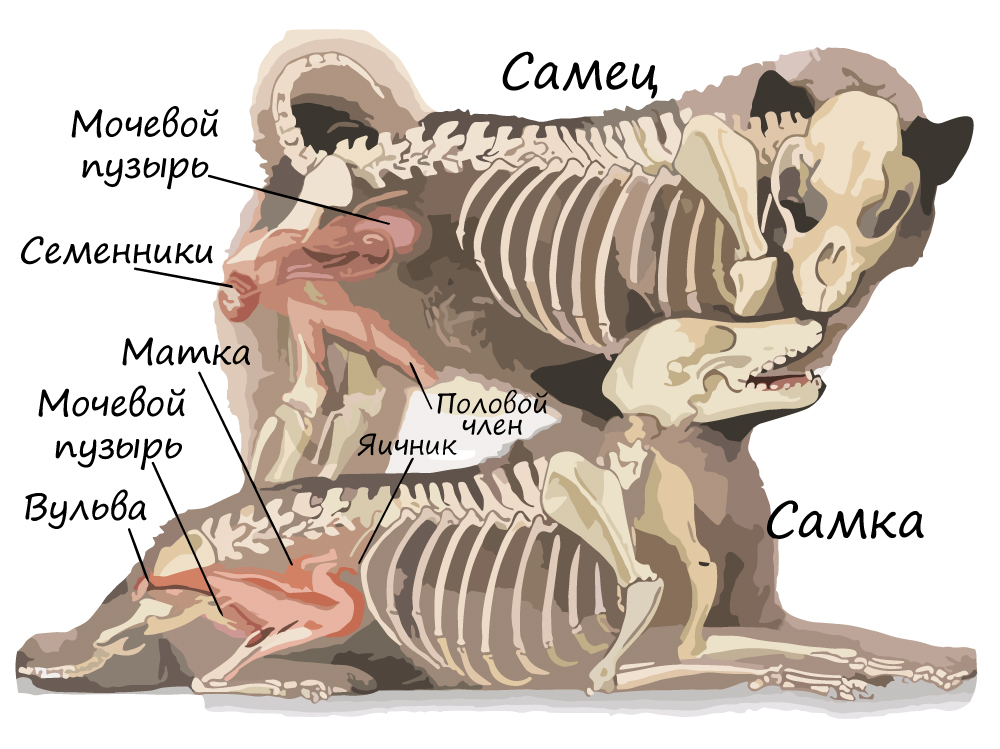
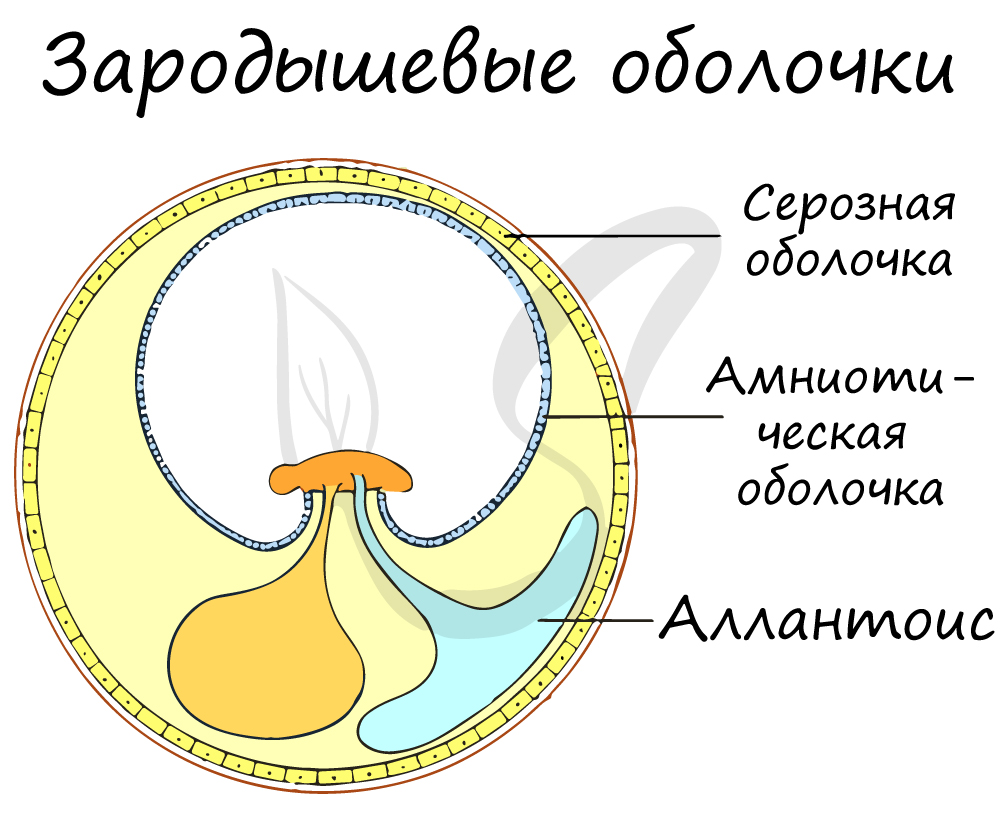
Значение млекопитающих

© Беллевич Юрий Сергеевич 2018-2021
Данная статья написана Беллевичем Юрием Сергеевичем и является его интеллектуальной собственностью. Копирование, распространение (в том числе путем копирования на другие сайты и ресурсы в Интернете) или любое иное использование информации и объектов без предварительного согласия правообладателя преследуется по закону. Для получения материалов статьи и разрешения их использования, обратитесь, пожалуйста, к Беллевичу Юрию.

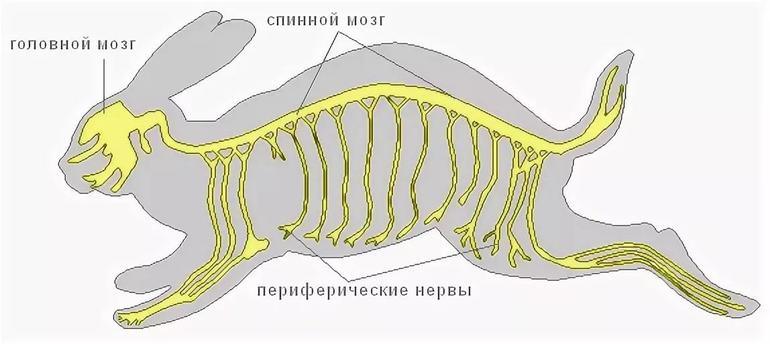
Отделы нервной системы
С учетом особенностей расположения органов нервной системы исследователи разделили ее на центральную и периферическую. К центральной относятся:
Существует также классификация, основанная на функциональных особенностях. По этим критериям нервную систему подразделяют на 2 вида:
Вегетативный отдел подразделяется на симпатический, парасимпатический и метасимпатический. Первые два подчиняются коре больших полушарий головного мозга и гипоталамусу. Метасимпатическая система представляет собой совокупность мелких нервных узлов и сплетений, которые расположены в стенках внутренних органов: мочевого пузыря, желудка, кишечника, сердца.
Импульсы по волокнам висцеральной системы движутся в обоих направлениях — от периферии к центру (афферентные) и обратно (эфферентные). На симпатический и парасимпатический отделы подразделяется эфферентная часть висцеральной системы.
Кроме функциональных имеются и анатомические различия. В частности, двигательные нейроны в соматической нервной системе располагаются в спинном мозге, а в вегетативной — за его пределами (в вегетативных нервных узлах).
Анатомия и физиология
В процессе онтогенеза нервная ткань млекопитающих развивается из эктодермального листка. Она состоит из нейронов (нервных клеток) и нейроглиальных клеток, несущих вспомогательную функцию.
Нейроны имеют множество отростков, функция которых заключается в проведении нервных импульсов, генерирующихся в клетке. Отростки нервных клеток образуют между собой многочисленные контакты (синапсы). Благодаря им импульс передается в нужном направлении. Эти отростки в совокупности с нейроглиальными клетками образуют белое вещество мозга. Нейроны сосредоточены в головном и спинном мозге.
Нервные волокна (нервы) представляют собой совокупность отростков нейронов, покрытых миелиновой оболочкой.
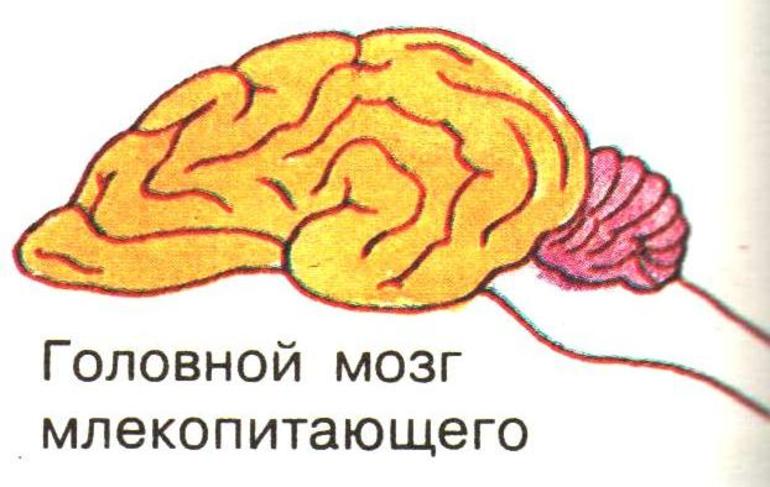
Головной и спинной мозг
Среди представителей животного мира именно у млекопитающих головной мозг наиболее развит. Это структуру можно разделить на следующие основные отделы:
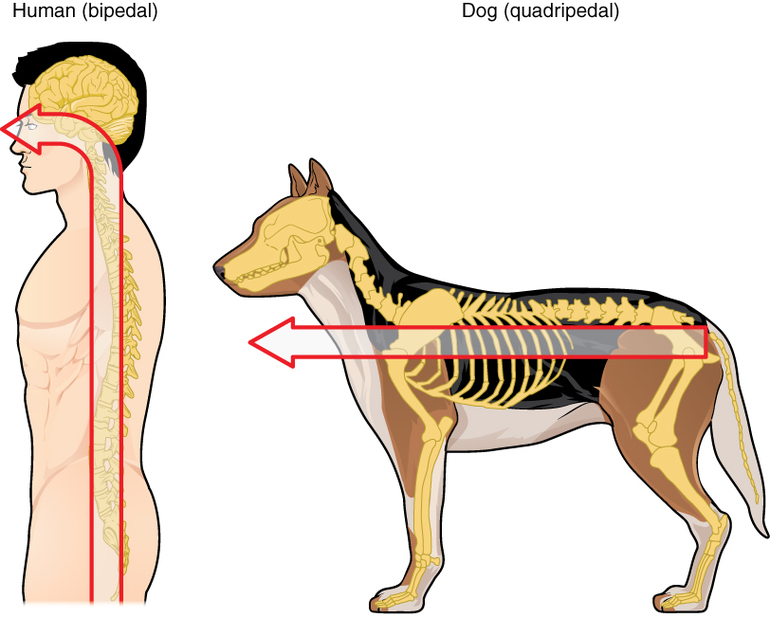
В спинном мозге серое вещество распределено неравномерно. На поперечном срезе оно образует фигуру, отдаленно напоминающую силуэт бабочки.
На таком срезе хорошо видны развитые восходящие пути, образованные афферентными центростремительными волокнами, передающими в головной мозг чувствительные импульсы. Также здесь есть нисходящие пути, сформированные двигательными центробежными волокнами, несущими импульсы от мозга к мускулатуре. Толщина этого органа неодинакова.
На уровне плечевого пояса и поясничной области он образует утолщения, выраженность которых зависит от степени развития соответствующих конечностей. У летучих мышей, например, больше развито плечевое, а у кенгуру — поясничное.
Периферические нервы
Периферическая нервная система млекопитающих образована черепными (12 пар) и спинномозговыми нервами. У человека их 31 пара (сколько их у других млекопитающих, зависит от конкретного вида).
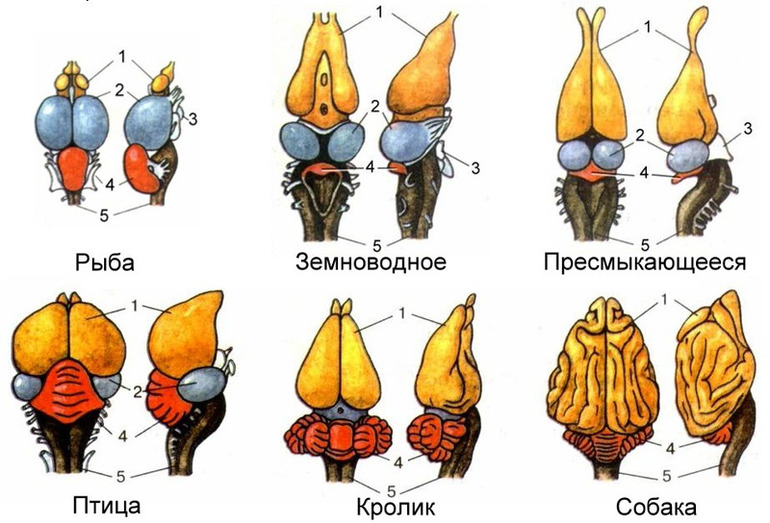
Каждая из них иннервирует соответствующий сегмент тела. В учебных пособиях по анатомии эта информация, как правило, кратко излагается в виде схемы или таблицы, где видно, какой корешок соответствует каждому сегменту.
Последние две пары черепных нервов относятся к соматической нервной системе, отвечая в том числе и за координацию.
Эта система хорошо развита у млекопитающих, что объясняет высокую скорость рефлексов.
Головной мозг (encephalon, или cerebrum) принимает на себя верховное командование функциями организма (за исключением самых простых рефлексов). В нем различается множество сложных, связанных друг с другом центров. Как было отмечено, внедрение ассоциативных нейронов в простую рефлекторную дугу сильно расширяет поле возможных вариантов ответа на данный сенсорный стимул и вместе с тем резко увеличивает разнообразие воздействий, которые могут вызвать определенный двигательный ответ. Работа головного мозга основана в значительной степени на этом принципе — посредничестве серии дополнительных нейронов между первичными чувствительными областями и конечными двигательными путями. Эти промежуточные нейроны сгруппированы в функциональных центрах. Такие центры могут соотносить и интегрировать афферентные импульсы для достижения адекватных ответов или координировать двигательные механизмы. На более высоких уровнях развиваются ассоциативные центры, которые существенно расширяют способности к исследовательскому поведению, обучению и накоплению индивидуального опыта. Этому способствует также и необычайное увеличение ресурсов оперативной, краткосрочной и долгосрочной памяти.
Первоначально волокна в головном мозге, примерно как и в спинном, пересекались во всех направлениях, соединяя самые разные области. В дальнейшем большинство нервных клеток, выполняющих одинаковые функции, объединились в центры, а волокна со сходными связями — в аккуратные, хотя зачастую с трудом обнаруживаемые пучки. Некоторые центры имеют специальные названия, однако большинство из них на-зываются ганглиями (узлами) или ядрами. Центры, или ядра, в широком смысле варьируют по размеру от микроскопических групп клеток, лежащих в сером веществе ствола мозга, до таких крупных структур, как кора мозжечка или переднего мозга. Пучки волокон, связывающие ядра одно с другим, обычно называются проводящими путями (трактами); волокна, образующие такой тракт, естественно, представляют собой аксоны нейронов, тела которых лежат в ядре, из которого он выходит. Проводящие пути обычно носят названия, состоящие из двух частей, которые указывают их начало и конец; так, кортикоспинальный тракт проводит импульсы из коры полушарий переднего мозга к двигательным нейронам спинного мозга (прямо или опосредованно). 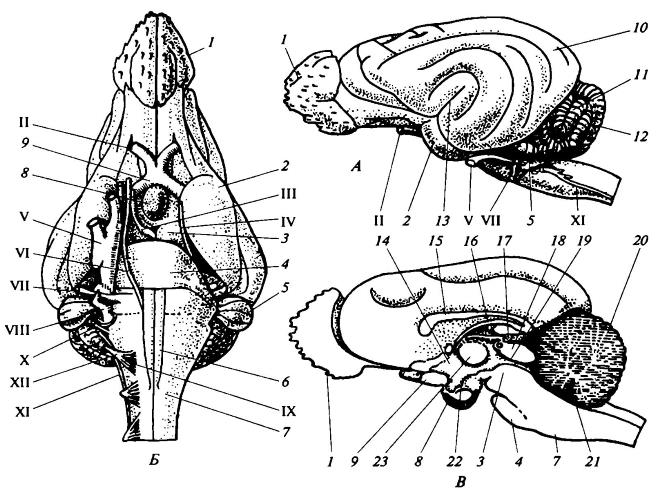
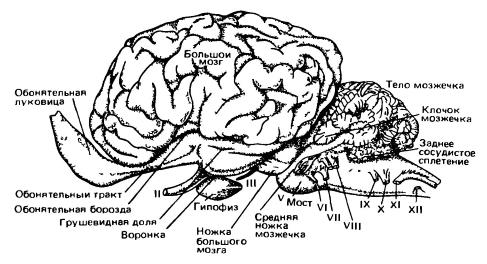
Головной мозг песца (Аlорех lagopus) (по Дзержинскому, 2013):
А — вид сбоку; Б — вил снизу; В — сагиттальный разрез; 1 — обонятельная луковица; 2 — древняя кора (грушевидная доля); 3 — средний мозг; 4 — мост; 5 — клочок мозжечка; 6 — пирамида; 7 — продолговатый мозг; 8 — гипофиз; 9 — перекрест зрительных нервов; 10 — полушарие переднего мозга; 11 — полушарие мозжечка; 12 — червяк мозжечка; 13 — сильвиева борозда; 14 — промежуточный мозг; 15 — мозолистое тело; 16 — свод; 17— эпифиз; 18 — область четверохолмия; 19 — мозговой водопровод; 20 — мозжечок; 21 — ромбовидная ямка; 22 — воронка; 21 — зрительный бугор; I-XII — черепные нервы. Несмотря на общую тенденцию клеток и волокон головного мозга собираться в четко ограниченные центры и пути, ретикулярная формация (formatio reticularis) сохраняет примитивное строение. Это образование представляет собой сплетение тел и волокон нейронов, связанное с двигательными столбами ствола мозга и передней части спинного мозга; особенно хорошо ретикулярная формация развита в передней части ствола мозга. К ней сходится множество чувствующих путей. Эта система используется для координации движений. Кроме того, ретикулярная формация участвует в передаче нисходящих сигналов из передних отделов головного мозга к двигательным центрам продолговатого и спинного мозга. Ретикулярная формация имеет у млекопитающих важное значение, не только сохраняя роль низшего звена пути к двигательным центрам, но также действуя в противоположном направлении как центр, стимулирующий высшие центры коры переднего мозга. Она также контролирует сложные рефлексы, такие, как чихание и кашель.
Головной мозг по своему строению билатерально симметричен; следовательно, во избежание «раздвоения личности» должны установиться перекрестные связи. Даже в спинном мозге имеется множество ассоциативных нейронов, волокна которых переходят на противоположную сторону. В головном мозге такие связи также многочисленны. Здесь имеется ряд комиссур (спаек) — проводящих путей, которые связывают одноименные области противоположных сторон. Кроме того, в некоторых случаях продольные проводящие пути переходят с одной стороны на другую (т. е. сворачивают под прямым углом) иногда без видимой причины. Такое явление уже описывалось для зрительных нервов (которые на самом деле представляют собой проводящие пути головного мозга). Другим примером служит тот факт, что движения частей одной стороны тела контролируются серым веществом полушария противоположной стороны — результат перекреста кортикоспинальных проводящих путей двух сторон. Система «проводки» головного мозга выше обсуждалась как система прямой передачи из одного конца в другой, от рецептора к эффектору, через цепь нейронов. Однако становится все очевиднее, что общая функциональная схема мозга гораздо сложнее и включает обратные связи, обходные пути, связанные с памятью и научением, и другие сложные механизмы. 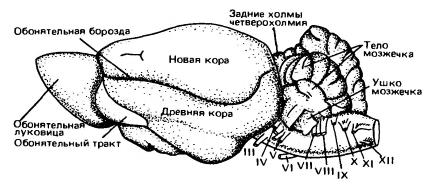
Мозг гимнуры (Echinosorex gymnura) (вверху) и лошади (Equus ferus) (внизу); вид сбоку. Римскими цифрами обозначены черепные нервы. (По Ромеру и Парсонсу, 1992.) 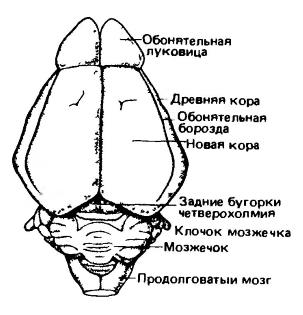
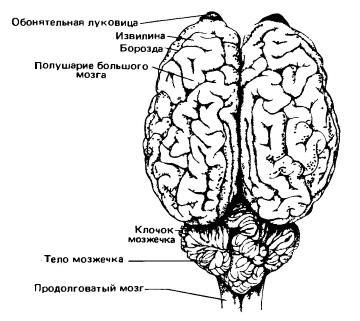
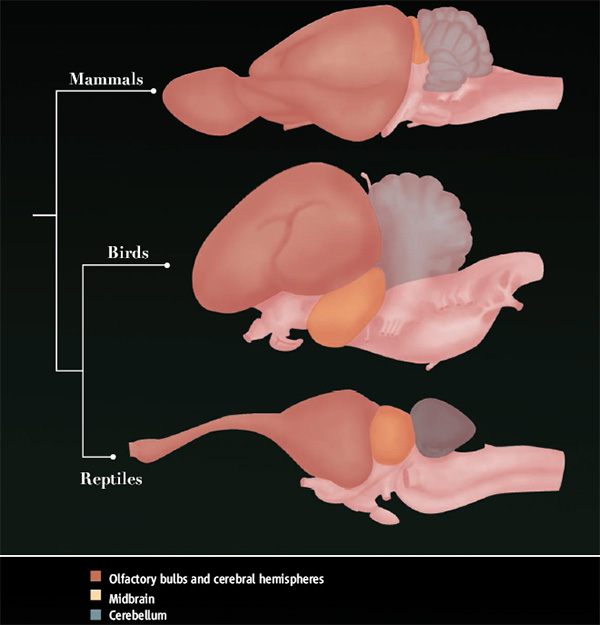
Мозг гимнуры (Echinosorex gymnura) (слева) и лошади (Equus ferus) (справа); вид сверху. (По Ромеру и Парсонсу, 1992.) Итак, головной мозг млекопитающих разделяется на несколько неравнозначных отделов — продолговатый мозг, мозжечок, средний мозг, промежуточный мозг и передний мозг. Каждый отдел обнаруживает специфические для класса особенности.
Продолговатый мозг (myelencephalon, или medulla oblongata) обладает чертами сходства со спинным мозгом, в который он переходит и с которым соединен мощными нервными трактами. Его центральный канал преобразуется в четвертый мозговой желудочек, в результате чего левые и правые столбы серого вещества сверху сильно разнесены. При переходе в спинномозговой канал желудочек формирует ромбовидную ямку (fossa rhomboidea). Продолговатый мозг утолщается не только вследствие расширения внутреннего канала, но также за счет наслоения снизу новых пучков продольных волокон (т. н. пирамид) и развития нейронных скоплений (олив). На большей части длины крыша этого мозгового отдела тонкая, перепончатая и складчатая; она формирует заднее сосудистое сплетение (plexus chorioideus posterior). Продолговатый мозг содержит ряд жизненно важных нервных центров (работы сердца, дыхания, пищеварения) и дает начало большинству черепных нервов (с VI-й по XII-ю пару).
Над передней частью продолговатого мозга возвышается хорошо развитый мозжечок (metencephalon, или cerebellum). Как и у птиц, он монолитен, т. е. лишен внутренней полости. В состав мозжечка входят непарный средний и парные боковые отделы. Средний отдел представляет собой первичную структуру мозжечка — его тело (corpus cerebelli), к которому подходят чувствующие нервные волокна. Боковые отделы взаимодействуют преимущественно с корой переднего мозга. Уровень их развития коррелирует с повышением роли коры в командовании двигательными функциями и развитием крупных нервных пучков, ведущих из двигательной коры в мозжечок и обратно. У однопроходных, как и у рептилий, тело мозжечка преобладает над его боковыми отделами, которые располагаются в виде незначительных придатков. У сумчатых, ксенартр, рукокрылых и грызунов боковые отделы по своим размерам не уступают телу. У хищных и копытных эти отделы становятся больше тела, образуя полушария мозжечка (hemispheria cerebelli). Наконец, у приматов тело на фоне сильно развитых полушарий выглядит совсем небольшим. Оба полушария связаны лежащей под ними комиссурой из поперечных нервных волокон. В стороны от полушарий мозжечка выступают небольшие булавовидные придатки — т. н. клочки (flocculi).
В отличие от всех прочих участков центральной нервной системы, не считая полушарий переднего мозга и крыши среднего мозга, в мозжечке содержащее нейроны серое вещество располагается на поверхности в виде слоистой коры, а белое вещество находится глубже. Образованный серым веществом поверхностный листок часто собран глубокими складками, благодаря чему он достигает большей площади. Центральный продольный ряд поперечных складок именуется червяком. Под серым веществом лежит белое, в котором веерообразно расходятся афферентные волокна, идущие во всех направлениях к поверхности, а также эфферентные волокна, идущие назад из серого вещества. Кора мозжечка содержит клетки нескольких хорошо различимых типов, расположенные слоями; с помощью сложного переплетения волокон между этими клетками осуществляется взаимный обмен сенсорными сигналами. Наиболее замечательные клеточные элементы — это клетки Пуркинье; их дендриты образуют густо ветвящиеся деревца, собирающие импульсы с обширного участка коры, и посылают эфферентные сигналы по своим аксонам в центральное ядро (nucleus centralis), где происходит переключение. В мозжечке наблюдается довольно четкая локализация функций, однако тело мозжечка действует во многих отношениях как интегрированный блок.
Пучки нервных волокон, ведущие к мозжечку и от него, образуют три пары столбообразных структур, называемых ножками мозжечка (pedunculi cerebelli). Передние ножки, состоящие главным образом из эфферентных пучков, соединяют мозжечок со средним мозгом. Средняя, или латеральная, пара ножек направлена из варолиевого моста вверх вдоль боковых сторон продолговатого мозга. В этих ножках сосредоточены волокна, идущие из коры переднего мозга в мозжечок; они перекрещиваются (пересекают среднюю линию) и переключаются в вентральной области моста. Задние ножки восходят из продолговатого мозга и заключают проприоцептивные волокна, идущие от спинного мозга. 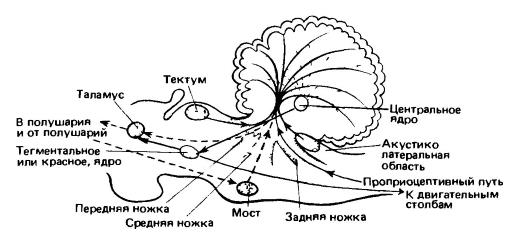
Схема основных связей мозжечка. Свойственные млекопитающим связи с корой мозжечка показаны штриховыми линиями. (По Ромеру и Парсонсу, 1992.) Мозжечок играет исключительно важную роль в поддержании равновесия тела и координации двигательной активности, отличающейся у млекопитающих большой сложностью. В его тело направляются волокна проприоцепторов сухожилий и мышечных веретен, фиксирующих положение частей тела и напряжение мышц, а также волокна равновесных рецепторов внутреннего уха, воспринимающих ускорение. В дополнение к этим двум первичным источникам сенсорная картина, складывающаяся в мозжечке, довершается некоторыми сигналами, приходящими по волокнам от чувствующих участков внешних покровов и зрительных центров. Далее, мозжечок снабжается информацией о мышечных движениях, направляемых высшими мозговыми центрами, но находящихся также под влиянием мозжечка. У млекопитающих, двигательные функции которых регулируются преимущественно корой переднего мозга, данные из него доставляются в мозжечок мощными пучками волокон через мост. Автоматически оценивая по полученным сигналам результаты предыдущих двигательных команд и коррелируя их с обобщенными сенсорными данными, мозжечок вырабатывает слабые тормозные импульсы, умеряющие интенсивность последующих команд. Импульсы отправляются по восходящим и нисходящим пучкам эфферентных волокон вдоль обеих сторон ствола мозга к боковым стенкам среднего мозга (область покрышки), откуда они продолжаются (частично после переключения) до таламуса и оттуда до коры больших полушарий. В результате аналитической работы мозжечка эффективность совершаемых животным движений значительно повышается. Установлено также, что мозжечок — по крайней мере, у человека — принимает участие в сложных когнитивных процессах. Согласно проведенным исследованиям, он помогает планировать сложное поведение, организовывать последовательность действий и даже участвует в речевой деятельности. 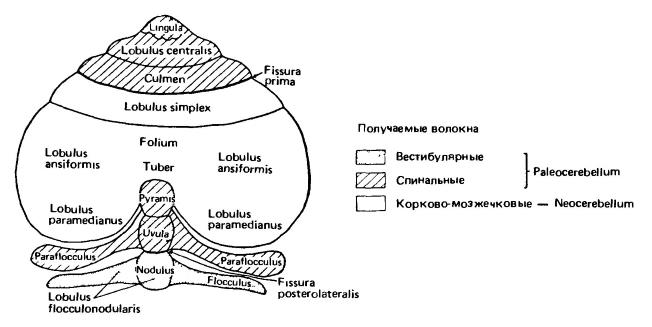
Схема внешнего строения мозжечка млекопитающего. Наиболее филогенетически древними отделами мозжечка являются отделы, связанные с чувством равновесия, — вестибулярные (обозначены точками), и отделы, связанные с мышечным чувством, — спинальные (выделены штриховкой). Белая зона представляет собой новоприобретение млекопитающих, связанное с корой полушарий большого мозга. (По Ромеру и Парсонсу, 1992.) Средний мозг (mesencephalon) имеет незначительные размеры. Желудочек этого мозгового отдела зажат его толстыми стенками в узкий канал, названный сильвиевым водопроводом. Лежащее выше желудочка серое вещество образует утолщение — тектум, или крышу (tectum). Более тонкие участки серого вещества в боковых стенках принято называть тегментумом, или покрышкой (tegmentum). Большую часть стенок и дна среднего мозга формируют волокнистые пучки белого вещества, в том числе крупные пучки двигательных волокон (пирамидные пути, нисходящие из полушарий), образующие ножки переднего мозга (crura cerebri).
У низших позвоночных и птиц крыша среднего мозга играет важную роль зрительного ассоциативного центра, принимая также стимулы из других сенсорных областей. У млекопитающих тектум претерпевает существенную редукцию, а большую часть его функций берет на себя серое вещество больших полушарий. Основные слуховые и прочие соматические сенсорные сигналы переключаются по пути в таламусе и далее идут в полушария. Лишь немногие из числа зрительных волокон продолжают следовать по своему первоначальному пути в средний мозг, основная же их масса прерывается в таламусе, и зрительные сигналы тоже переправляются вперед, к полушариям. И все-таки, крыша среднего мозга млекопитающих в определенной степени продолжает исполнять роль центра зрительных и слуховых рефлексов. Две взаимно перпендикулярные борозды разделяют тектум на четыре небольших вздутия, называемых четверохолмием (corpora quadrigemina). Передняя пара вздутий — верхние бугры (colliculi superiores) — соответствует зрительным долям низших позвоночных и осуществляет зрительные рефлексы. Задняя пара бугров (colliculi inferiores) представляет собой преобразованные слуховые центры, которые играют роль передающего центра для слуховых сигналов на их пути в таламус, а затем и в большие полушария. У некоторых групп млекопитающих задние холмы достигают значительных размеров.
Область стенки среднего мозга, называемая покрышкой, в основном представляет собой переднее продолжение двигательных областей продолговатого мозга. В связи с этим она функционирует как отдел, в котором различные импульсы из промежуточного мозга, тектума и мозжечка координируются и передаются вниз к двигательным ядрам ствола мозга и спинного мозга. Помимо волокон ретикулярной системы, в покрышке имеется т. н. красное ядро (nucleus ruber), где осуществляется переключение эфферентных сигналов из мозжечка.
Варолиевый мост (pons Varolii) соединяет у млекопитающих средний мозг, продолговатый мозг и мозжечок. Внешне он имеет вид поперечно утолщенного валика из белого нервного вещества, перекрывающего снизу переднюю часть продолговатого мозга. Внутри него проходит множество проводящих путей, связывающих кору головного мозга со спинным мозгом и с корой полушарий мозжечка; кроме того, здесь находится несколько ядер серого вещества. От передней поверхности моста отходит V-я пара нервов (тройничные). 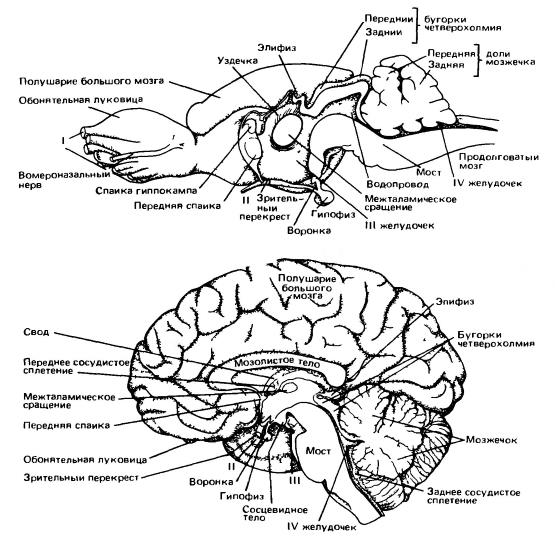
Дорсальная и вентральная части промежуточного мозга, его крыша и дно, именуются эпиталамус (epithalamus) и гипоталамус (hypothalamus). Почти на всем своем протяжении эпиталамус имеет незначительную толщину и не содержит нейронов. Спереди этой тонкой крыши располагается переднее сосудистое сплетение (plexus chorioideus anterior), втянутое в полости желудочков (непарного и боковых). Дальше назад находится шишковидное тело, или эпифиз (epiphysis cerebri). У низших позвоночных этот небольшой орган сообщается с внешней средой через непарное теменное отверстие и именуется теменным глазом. Он содержит фоторецепторные клетки, которые воспринимают интенсивность света, но не способны давать изображение. В эпифизе млекопитающих фотопигменты в существенном количестве не синтезируются и наружные структуры, обеспечивающие светочувствительность, отсутствуют; они обнаружены лишь у новорожденных некоторых видов. Полость эпифиза у взрослых зверей также исчезает, единственное известное исключение из этого правила составляет виргинский опоссум (Didelphis virginiana). Информацию о том, светло или темно, эпифиз взрослых млекопитающих получает по нервам из таламуса. Кроме того, эпифиз является важной железой внутренней секреции.
В передней части гипоталамуса располагается очень заметное образование — зрительный перекрест (chiasma opticum). Дно третьего желудочка составляет воронка (infundibulum). Ниже находится тесно соединенный с воронкой гипофиз (hypophysis cerebri), являющийся важнейшим элементом эндокринной системы. Он участвует в сезонной перестройке метаболизма и регуляции периодических явлений — линьки, спячки, размножения, миграций и др.
В гипоталамусе располагаются обонятельные центры, среди которых заслуживают упоминания сосцевидные тела (corpora mamillaria), соседствующие с гипофизарной областью. Однако основная функция гипоталамуса — роль главного вегетативного центра (другие важнейшие центры носят почти исключительно соматический характер). Многие вегетативные нервные функции выполняются за счет рефлексов спинного или продолговатого мозга. В свою очередь, гипоталамус служит важнейшим интегративным центром вегетативной деятельности, располагая прямыми двигательными связями с вегетативными центрами мозгового ствола и спинного мозга. В различные ядра этой области поступают сигналы от органов обоняния и вкуса, а также сенсорные импульсы из всевозможных вегетативных структур. Особенно важное значение имеют связи с чисто обонятельными участками больших полушарий; восприятие запахов играет огромную роль в активности вегетативных отделов нервной системы. Нисходящие пути связывают ядра с лежащими позади них центрами вегетативной нервной системы.
Диапазон регуляторных функций гипоталамуса выяснен пока далеко не полностью. Известно, однако, что здесь находится один из чувствительных «термостатов», обеспечивающий регуляцию температуры тела. Частота сердечных сокращений, интенсивность дыхания, кровяное давление, сон и деятельность кишечника также регулируются гипоталамусом или, во всяком случае, находятся под его влиянием. Кроме того, гипоталамус вовлечен в регуляцию деятельности гипофиза. Некоторые его гормоны в действительности синтезируются гипоталамическими ядрами, откуда по кровеносным сосудам передаются в нервную долю гипофиза.
Продолговатый мозг, средний мозг и варолиев мост традиционно объединяются в мозговой ствол; иногда в него включают и промежуточный мозг. В сером веществе продолговатого мозга и задней части среднего мозга имеется серия столбов, в принципе сходных с таковыми спинного мозга. И там, и тут присутствует спинной столб, содержащий ассоциативные нейроны, которые получают и распределяют сенсорные импульсы, и брюшной столб, содержащий двигательные нейроны. Оба столба подразделяются на соматический и вегетативный компоненты. Главное отличие от спинного мозга — сильное расширение центрального канала. С каждой стороны по внутренней поверхности ствола мозга проходит горизонтальная пограничная борозда (sulcus limitans), которая отделяет дорсальную чувствующую зону от вентральной двигательной.
Столбы серого вещества мозгового ствола распадаются на отдельные ядра, которые соответствуют ответвлениям черепных нервов и являются центрами различных функций организма, контролируемых стволом мозга или находящихся под его влиянием. Соматически-двигательный столб разделен на несколько расположенных в среднем мозге и в переднем конце продолговатого мозга мелких ядер нервов глазных мышц и более заднее ядро подъязычного нерва. Вегетативно-двигательный столб разбит на отдельные особые вегетативно-двигательные ядра V-й, VII-й и IX-й+X-й пар нервов (последним двум принадлежит двойное ядро (nucleus ambiguus)). В этом столбе также образуется дыхательный центр, участвующий в регуляции ритмики дыхания. Также имеются небольшие парасимпатические ядра: в среднем мозге, отвечающее за глазные рефлексы; в передней части продолговатого мозга, обслуживающее слюнные железы; в задней части продолговатого мозга, принадлежащее соответствующему компоненту блуждающего нерва. Ядра разных нервов, связанные с обычными вегетативно-чувствующими волокнами, объединены в одиночное ядро (nucleus solitarius) продолговатого мозга, но параллельно имеется особое вегетативно-чувствующее, или вкусовое, ядро (nucleus gustatorius). Соматически-чувствующий столб является, по сути, единым длинным ядром, которое главным образом связано с тройничным нервом и тянется на протяжении большей части длины ствола мозга, заходя в спинной мозг. 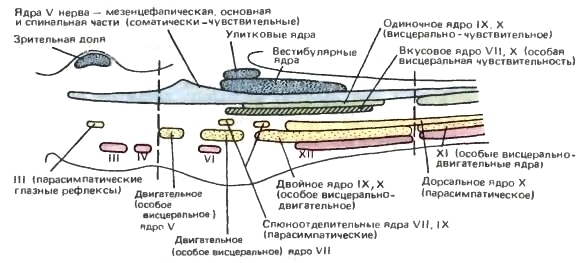
Схемы областей среднего и заднего мозга млекопитающих; вид сбоку. Показано расположение чувствующих и двигательных ядер. Соматически-чувствующие ядра обозначены голубым цветом; особые соматически-чувствующие ядра выделены точками; вегетативно-чувствующие — окрашены в зеленый цвет; особые вегетативно-чувствующие — заштрихованы; вегетативно-двигательные обозначены желтым; особые вегетативно-двигательные выделены точками; соматически-двигательные выделены красным. Соматически-чувствующий столб в целом сохраняет непрерывность (он связан практически исключительно с V-м нервом), но прочие столбы разбиты на отдельные ядра. Вегетативно-чувствующий столб включает регулярное вегетативное ядро (преимущественно для афферентных волокон, которые приходят от внутренних органов в составе блуждающего нерва) и особое ядро для восприятия одного из важных чувств — вкуса. Эфферентные вегетативные ядра представлены мелкими передними центрами, осуществляющими автономные глазные рефлексы и контролирующими слюнные железы, а также крупным ядром, откуда в составе блуждающего нерва парасимпатические волокна направляются к внутренностям. Важную роль играют вегетативно-двигательные ядра, связанные с V-м иVI-м, а также с IX-м и X-м нервами. Соматически-двигательный столб включает мелкие ядра для глазных мышц спереди и крупное подъязычное ядро позади. (По Ромеру и Парсонсу, 1992.) Серое вещество мозгового ствола покрыто толстым слоем белого вещества, представляющего собой проводящие пути между спинным мозгом и более передними центрами головного мозга. Подобно серому, белое вещество в стволе организовано примерно так же, как и в спинном мозге, но вдобавок большое число восходящих и нисходящих волокон переходит в продолговатом мозге на противоположную сторону.
Системы двигательных и чувствующих столбов и ядер продолговатого мозга достаточно для построения рефлекторных дуг между рецепторами и обеспечивающими ответ исполнительными органами головы. Однако в головном мозге также действуют и более сложные механизмы. Вдобавок к простым рецепторам кожи, мышц и кишечника, имеющимся во всем теле, в голове присутствуют особые органы чувств. Соответственно должны существовать центры, воспринимающие первичные сигналы от этих органов, и центры более высокого уровня, соотносящие и интегрирующие эти сигналы, прежде чем будут даны окончательные руководящие указания двигательным столбам ствола мозга и спинного мозга. Большинство таких центров расположено в других отделах головного мозга. Однако даже в относительно просто организованном продолговатом мозге обнаруживается первичная чувствующая зона одной из главных сенсорных систем — акустической. Она представлена слуховыми ядрами, которые находятся в передней части продолговатого мозга и имеют отдельные центры для преддверия и для улитки внутреннего уха.
В отличие от задней части мозгового ствола, средний и промежуточный отделы обнаруживают специализированные функциональные черты. Из продолговатого мозга в средний на некоторое расстояние продолжаются ряды чувствующих и двигательных ядер, связанных с черепными нервами. Однако дальше впереди подобные структуры отсутствуют. Передние центры ствола мозга, вообще говоря, не имеют прямой связи с афферентными или эфферентными сигналами, не считая импульсов зрительного нерва. Эта область выполняет две основные функции. Во-первых, она представляет собой основную станцию на пути между нижележащими участками мозга и большими полушариями. Во-вторых, она служит важным местом расположения центров нервной корреляции и координации. Впрочем, у млекопитающих функции последнего типа на фоне деятельности коры больших полушарий заметны слабо. 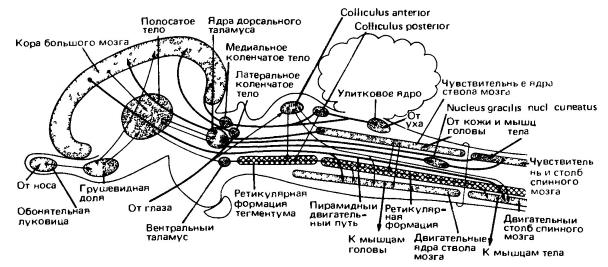
Упрощенная схема проводящих путей в мозге млекопитающего. Роль крыши среднего мозга (tectum) ограничена, он становится второстепенным рефлекторным центром. Полосатое тело (corpus striatum) имеет относительно небольшое значение. Большинство сенсорных импульсов направляется «вверх» в кору полушарий переднего мозга, откуда прямой двигательный путь (пирамидный путь) направляется к двигательным центрам ствола мозга и спинного мозга. (По Ромеру и Парсонсу, 1992.) вперед



