В какие отделы мозга поступает информация с проприорецепторов
Проприоцептивные ощущения дают человеку возможность воспринимать изменения положения отдельных частей тела в покое и во время совершаемых движений. Информация, поступающая от проприоцепторов, позволяет ему постоянно контролировать позу и точность произвольных движений, дозировать силу мышечных сокращений при противодействии внешнему сопротивлению, например при подъеме или перемещении груза.
Проприоцепторы представлены интрафузальными волокнами мышц (датчики длины мышцы), сухожильными рецепторами Гольджи (датчики напряжения мышцы) и механорецепторами суставных капсул, отвечающими изменениями фоновой активности на сгибание или разгибание в суставе и морфологически сходными с тельцами Руффини и рецепторами Гольджи.
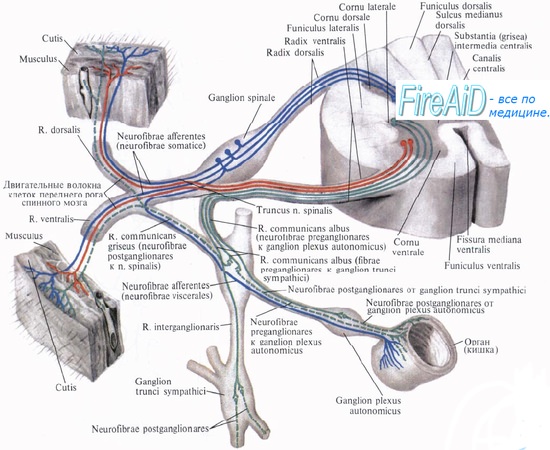
Афферентная информация от проприоцепторов в составе задних корешков поступает в спинной мозг и без переключения в нем передается по восходящим путям к ядрам задних столбов продолговатого мозга. Расположенные там сенсорные нейроны второго порядка передают информацию по лемнисковому пути на противоположную сторону мозга к проекционным ядрам таламуса. Нейроны переключательных ядер таламуса образуют соматотопическую проекцию на соматосенсорную кору, с помощью которой формируются проприоцептивные ощущения.
Сенсорные сигналы от проприоцепторов мышц лица и головы следуют в ЦНС по чувствительным волокнам черепных нервов, а в соматосенсорную кору информация поступает тем же способом, что и сигналы от мускулатуры тела, т. е. по лемнисковому пути.
Проприоцептивное восприятие положения тела и движений происходит в результате объединения в соматосенсорной коре информации от всех разновидностей проприоцепторов. Порог воспринимаемых изменений углового положения проксимальных суставов ниже, чем дистальных: 0,2— 0,4 градуса в плечевом суставе и около 1 градуса в суставах пальца руки. Порог возрастает при увеличении угловой скорости движений, в связи с чем уменьшается точность выполняемых движений. Способность человека дифференцировать силу произвольных сокращений мышц подчиняется закону Вебера— Фехнера, эта способность уменьшается при самых слабых и очень сильных мышечных усилиях, необходимых для выполнения работы.
В какие отделы мозга поступает информация с проприорецепторов
Проприоцепция — это группа сигналов, посылаемых в центральную нервную систему специальными терминалами (проприоцепторами), расположенными в суставных капсулах, связках, сухожилиях и мышцах.
Проприоцепторы, среди которых выделяют мышечные рецептор ных образований» href=»http://www.braintools.ru/article/9580″> рецептор ы, или мышечные веретена ( рецептор ы растяжения), сухожильные рецептор ы, или органы Гольджи ( рецептор ы мышечной силы), а также суставные рецепторы относятся к механо рецептор ам, посылающим в ЦНС информацию о положении, деформации и смещениях различных частей тела.
Функционирование этих рецептор ов обеспечивает координацию всех подвижных органов и тканей животного и человека в состоянии покоя и во время любых двигательных актов. При экспериментальном выключении проприоцепторов животные теряют способность поддерживать естественные позы, двигаться и целесообразно реагировать на внешние воздействия.
Если человек закроет глаза и попытается написать текст, то буквы будут написаны достаточно четко. Этим простым способом легко убедиться в умении человека пользоваться информацией, идущей от мышц и суставов.
Двигательный, или кинестетический, анализатор (мышечная сенсорная система ) обеспечивает формирование так называемого мышечного чувства при изменении напряжения мышц, суставных сумок, связок и сухожилий. Проблема мышечно-суставных ощущений имеет исключительное значение для физиологии и психологии. Специфические особенности человека появились благодаря более совершенной организации мышечной сенсорной системы по сравнению с животными.
И.М. Сеченов считал, что мышечное чувство является ближайшим регулятором движений и одним из орудий ориентации человека в пространстве и времени. Ощущения положения и перемещения тела в пространстве, ощущения во время трудовой деятельности и членораздельная речь лежат в основе формирования сознания человека и его представлений об окружающем мире.
Мышечное чувство обладает тремя качествами. Это, во-первых, ощущение положения конечностей, когда человек может определить положение своих конечностей и их частей относительно друг друга. Во-вторых, ощущение движения, когда, изменяя угол сгибания в суставе, человек осознаёт скорость и направление движения. Третьим качеством является ощущение усилия, когда человек может оценить мышечную силу, нужную для движения или удерживания суставов в определённом положении при подъёме или перемещении груза.
Наряду с кожной, зрительной и вестибулярной сенсорными системами двигательный анализатор оценивает положение тела в пространстве, позу, участвует в координации мышечной деятельности.
Двигательная сенсорная система служит для анализа состояния двигательного аппарата — его движения и положения. Информация о степени сокращения скелетных мышц, натяжении сухожилий, изменении суставных углов необходима для регуляции двигательных актов и поз.
Общий план организации. Двигательная сенсорная система состоит из следующих 3-х отделов:
Функции проприо рецептор ов
К проприо рецептор ам относятся мышечные веретена, сухожильные органы (или органы Гольджи) и суставные рецептор ы ( рецептор ы суставной капсулы и суставных связок). Все эти рецептор ы представляют собой механо рецептор ы, специфическим раздражителем которых является их растяжение.
Мышечные веретена человека, представляют собой продолговатые образования длиной несколько миллиметров, шириной десятые доли миллиметра, которые расположены в толще мышцы. В разных скелетных мышцах число веретен на1 г ткани варьирует от нескольких единиц до сотни.
Сухожильные органы ( рецептор ы) Гольджи человека располагаются в зоне соединения мышечных волокон с сухожилием, последовательно по отношению к мышечным волокнам.
В результате последовательного крепления сухожильных органов к мышечным волокнам (а в ряде случаев — к мышечным веретенам), растяжение сухожильных механо рецептор ов происходит при напряжении мышц. Таким образом, в отличие от мышечных веретен, сухожильные рецептор ы информируют нервные центры о степени напряжения мыши, и скорости его развития.
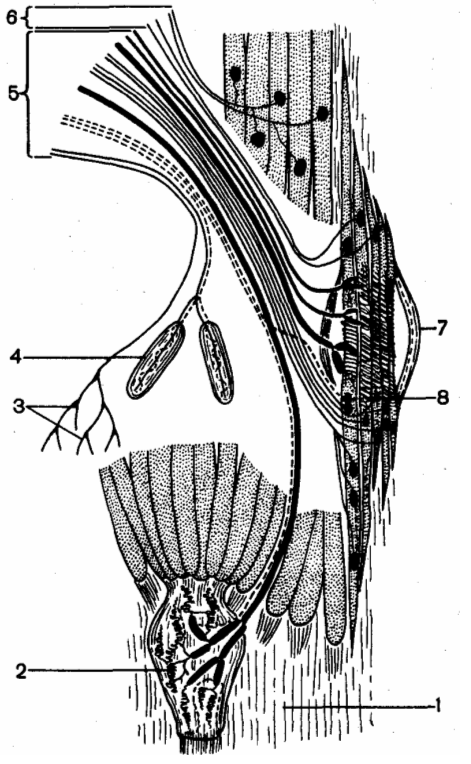
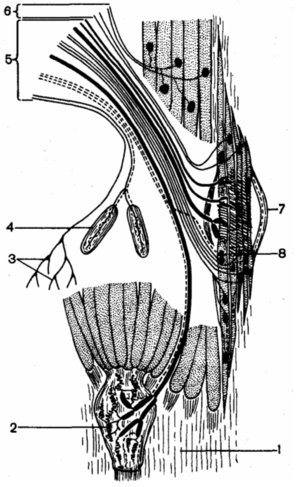
Эфферентная иннервация скелетной мышцы: 1 — сухожилие мышцы; 2 — сухожильные рецептор ы Гольджи; 3 — свободные окончания; 4 — тельца Фатера-Пачини; 5 — афферентные волокна; 6 — эфферентные волокна; 7 — мышечное веретено; 8 — окончания афферентных волокон
Суставные рецептор ы реагируют на положение сустава и на изменения суставного угла, участвуя, таким образом, в системе обратных связей от двигательного аппарата и в управлении им. Суставные рецептор ы информируют о положении отдельных частей тела в пространстве и относительно друг друга. Эти рецептор ы представляют собой свободные нервные окончания или окончания, заключенные в специальную капсулу. Одни суставные рецептор ы посылают информацию о величине суставного угла, т. е. о положении сустава. Их импульсация продолжается в течение всего периода сохранения данного угла. Она тем большей частоты, чем больше сдвиг угла. Другие суставные рецептор ы возбуждаются только в момент движения в суставе, т. е. посылают информацию о скорости движения. Частота их импульсации возрастаете увеличением скорости изменения суставного угла.
Аксоны таламических нейрон ов ( нейрон ов третьего порядка) оканчиваются в коре больших полушарий, главным образом, в соматосенсорной коре (постцентральная извилина) и в области сильвие-вой борозды (соответственно, участки S-1 и S-2), а также частично в двигательной (префронтальной) области коры. Эта информация используется двигательными системами мозга достаточно широко, в том числе для принятия решения о замысле движения, а также для его реализации. Кроме того, у человека на основе про-приоцептивной информации формируются представления о состоянии мышц и суставов, а также, в целом, о положении тела в пространстве.
Сигналы, идущие от рецептор ов мышечных веретен, сухожильных органов, суставных сумок и тактильных рецептор ов кожи. Механо рецептор ная и температурная чувствительность» href=»http://www.braintools.ru/article/9885″> рецептор ов кожи, называют кинестетическими, т. е. информирующими о движении тела. Их участие в произвольной регуляции движений различно. Сигналы от суставных рецептор ов вызывают заметную реакцию в коре больших полушарий и хорошо осознаются. Благодаря им человек лучше воспринимает различия при движениях в суставах, чем различия в степени напряжения мышц при статических положениях или поддержании веса. Сигналы же от других проприо рецептор ов, поступающие преимущественно в мозжечок, обеспечивают бессознательную регуляцию, подсознательный контроль движений и поз.
Таким, образом, проприоцептивные ощущения дают человеку возможность воспринимать изменения положения отдельных частей тела в покое и во время совершаемых движений. Информация, поступающая от проприоцепторов, позволяет ему постоянно контролировать позу и точность произвольных движений, дозировать силу мышечных сокращений при противодействии внешнему сопротивлению, например при подъеме или перемещении груза.
Проприоцептивное восприятие положения тела и движений происходит в результате объединения в соматосенсорной коре информации от всех разновидностей проприоцепторов.
Структура и функции проприорецепторов
В активной части опорно двигательного аппарата имеются проприорецепторы. Существует два типа проприорецепторов – нервно-мышечные веретёна, которые реагируют на изменение длины мышцы, а также сухожильные органы Гольджи, реагирующие на изменение напряжения, то есть на силу сокращения мышечных волокон (Рисунок 1).

Рисунок 1. Проприорецепторы: нервно-мышечное веретено и сухожильный орган Гольджи
Нервно-мышечные веретёна. Нервно-мышечные веретена расположены в толще скелетных мышц между мышечными волокнами. Они представляют собой веретеновидные структуры, покрытые соединительнотканной капсулой. Длина веретена составляет 4 – 7 (до 10) мм, диаметр – 100 мкм. Под капсулой внутри веретена находятся видоизменённые мышечные волокна, которые называются интрафузальными. Эти волокна имеют ряд отличий от обычных, экстрафузальных, мышечных волокон. Во-первых, интрафузальное волокно имеет меньший диаметр (15 – 30 мкм) по сравнению с экстрафузальным (50 – 100 мкм). Во-вторых, в центральной («экваториальной») части интрафузального волокна нет сократительных элементов, поэтому к сокращению способны только периферические части. Поскольку концы интрафузального волокна фиксированы к соединительнотканной капсуле, покрывающей веретено, при укорочении периферических отделов волокна его центральная часть растягивается. Интрафузальные волокна расположены параллельно эктсрафузальным. По строению интрафузальные волокна делятся на два типа – с ядерной сумкой и с ядерной цепочкой (Рисунок 2). В свою очередь, волокна с ядерной сумкой по функции по особенностям иннервации подразделяются на статические и динамические. Для волокон с ядерной сумкой характерно наличие расширенной центральной части, в которой ядра собраны в скопления. Волокна с ядерной цепочкой примерно вдвое тоньше волокон с ядерной сумкой, ядра в их центральной части расположены в ряд. В каждом веретене содержится 1 – 3 волокна с ядерной сумкой и 5 – 7 волокон с ядерной цепочкой.

Рисунок 2. Нервно-мышечное веретено
Интрафузальные волокна получают как афферентную, так и эфферентную иннервацию. Выделяют два типа афферентных окончаний – первичные и вторичные. Первичные окончания называют аннулоспиральными, они представляют собой спираль, окружающую центральную часть всех интрафузальных волокон. От первичных окончаний отходят волокна типа Ia, которые соответствуют типу Aα по Гассеру-Эрлангеру. Это самые толстые волокна (
17 мкм) с самой высокой скоростью проведения возбуждения (70 – 12 м/с). Каждое нервно-мышечное веретено иннервируется только одним волокном Ia. Другими словами одно афферентное нервное волокно Ia получает импульсацию со всех первичных окончаний в иннервируемом им веретене. Вторичные окончания имеют форму спирали, но могут также образовывать разветвлённую структуру. Они оплетают центральную часть интрафузальных волокон с ядерной цепочкой, а также статических волокон с ядерной сумкой, но их не бывает в динамических волокнах с ядерной сумкой. От вторичных окончаний отходят волокна типа II, соответствующие типу Аβ по Гассеру-Эрлангеру; диаметр этих волокон составляет
8 мкм, они проводят возбуждение со скоростью 40 – 70 м/с (Рисунок 3).

Рисунок 3. Схема нервно-мышечного веретена
Как первичные, так и вторичные нервные окончания чувствительны к изменению длины интрафузальных волокон, однако характер ответной реакции различается. При медленном растяжении центральной части интрафузальных волокон импульсация появляется как в первичных, так и во вторичных рецепторах. Частота импульсов пропорциональная степени растяжения волокна. Такой ответ афферентных окончаний называют статическим. Однако при быстром внезапном растяжении волокна в первичных (но не вторичных) окончаниях отмечается мощный всплеск импульсации – динамический ответ. Следовательно, волокна Ia передают информацию как об абсолютной длине интрафузальных волокон (то есть о степени растяжения), так о скорости её изменения, тогда как волокна II – только об абсолютной длине. Следует понимать, что растяжение центральной части интрафузальных волокон может быть связано как с удлинением всей мышцы (интрафузальные волокна расположены параллельно экстарфузальным), так и при сокращении периферических отделов волокна (Рисунок 3).

Рисунок 4. Статический и динамический ответы рецепторов нервно-мышечных веретён
Эфферентная иннервация интрафузальных волокон представлена нервными волокнами типа Aγ, которые являются аксонами γ-мотонейронов спинного мозга. Диаметр этих волокон составляет
5 мкм, скорость проведения возбуждения в них – 15 – 40 м/с. Окончания Aγ волокон подходят к периферическим отделам интрафузальных волокон, при возбуждении γ-мотонейронов концевые участки интрафузальных волокон сокращаются, в результате чего их центральные отделы растягиваются. Выделяют γ-динамические и γстатические волокна, γ-динамические волокна иннервируют динамичекие волокна с ядерной сумкой, их активация повышает динамическую чувствительность окончаний Iа типа.
γ-статические волокна иннервируют волокна с ядерной цепочкой и статические волокна с ядерной сумкой, их возбуждение повышает уровень активности как первичных, так и вторичных окончаний (см. ниже).
Миотатический рефлекс (рефлекс растяжения)
В ответ на увеличение длины мышцы рецепторы нервно-мышечных веретён возбуждаются, и импульсация по волокнам Ia (и при быстром, и при медленном растяжении) или II (при медленном растяжении) поступает в спинной мозг. Войдя в серое вещество спинного мозга, волокно афферентного нейрона разветвляется и образует возбуждающие синапсы практически со всеми α-моторными нейронами, иннервирующими гомонимную (то есть эту же) мышцу. Возбуждение α-моторных нейронов вызывает сокращение экстрафузальных волокон, и мышца укорачивается. Следует отметить, что коллатерали афферентных нейронов переключаются также на вставочные тормозные нейроны, которые угнетают α-мотонейроны мышц антагонистов. Таким образом, растяжение мышцы вызывает её рефлекторное укорочение, при этом мышцы антагонисты расслабляются. Описанный рефлекс с рецепторов нервно-мышечных веретён называется миотатическим, или рефлексом на растяжение (Рисунок 4). Миотатический рефлекс является моносинаптическим, поэтому имеет очень небольшой латентный период (

Рисунок 5. Миотатический рефлекс с нервно-мышечных веретён
Различают фазический (динамический) и тонический миотатический рефлекс. Фазический рефлекс запускается с первичных нервных окончаний при быстром растяжении мышцы, например, при ударе по сухожилию неврологическим молоточком или при появлении внезапного препятствия совершаемому движению. Тонический рефлекс запускается с вторичных нервных окончаний при медленном (или постоянном) растяжении мышечных волокон; он имеет значение для поддержания вертикальной позы. Когда тело отклоняется от правильного положения, мышцы, участвующие в поддержании позы, растягиваются. В результате возбуждаются как первичные, так и вторичные окончания нервно-мышечных веретён, запускается миотатический рефлекс, и мышцы сокращаются; за счёт их сокращения тело возвращается в правильное положение. Когда у длительно стоящего человека (например, солдата в стойке «смирно») из-за утомления сгибаются ноги в коленных суставах, проприорецепторы четырёхглавых мышц реагируют на растяжение, и ноги снова выпрямляются. Постоянное стимулирующее влияние на α-мотонейроны со стороны афферентных волокон Ia и II типов вносит вклад в поддержание мышечного тонуса.
Функции γ-моторных нейронов. Понятие о γ-петле.
Как уже упоминалось, интрафузальные волокна получают эфферентную иннервацию со стороны γ-моторных нейронов. При возбуждении γ-мотонейронов происходит сокращение периферических (концевых) отделов интрафузальных волокон, в результате их центральная часть растягивается. При этом первичные и вторичные нервные окончания активируются, импульсация от них поступает в спинной мозг, и возбуждаются α-моторные нейроны. Другими словами, запускается миотатический рефлекс. Такой механизм (активация рефлекса растяжения при возбуждении γ-мотонейронов) называется γ-петлёй. Наличие γ-петли обеспечивает сохранение чувствительности рецепторов нервномышечных веретён при разной длине мышцы. Во время сокращения мышцы её длина уменьшается, соответственно должны укорачиваться и интрафузальные волокна. В результате импульсация с рецепторов нервно-мышечных веретён должна была бы прекратиться, и рецепторы потеряли бы способность реагировать на изменение длины мышцы. Напротив, при расслаблении мышцы её длина увеличивается, и интрафузальные волокна могут быть чрезмерно растянуты. В таких условиях рецепторы будут генерировать импульсы с максимальной частотой и, следовательно, также потеряют способность реагировать на изменение длины. Однако в реальных условиях имеет место α-γ-коактивация, то есть α- и γмотонейроны возбуждаются одновременно. В результате сокращаются как экстра-, так и интрафузальные волокна. Сокращение экстрафузальных волокон приводит к укорочению мышцы, а сокращение периферических отделов интрафузальных волокон – к растяжению их центральной части. Следовательно, длина центральной части не меняется, поскольку её укорочение вследствие сокращения экстрафузальных волокон компенсируется растяжением из-за сокращения периферических отделов интрафузальных волокон. И наоборот – расслабление всей мышцы сопровождается расслаблением и интрафузальных волокон, при этом растяжение центральной части уменьшается. Таким образом, за счёт α-γ-коактивации рецепторы нервно-мышечных веретён сохраняют чувствительность (и, следовательно, передают информацию о длине мышцы в ЦНС) как при сокращении мышцы, так и при её расслаблении (Рисунок 6). На γ-мотонейронах заканчиваются нисходящие пути из различных отделов головного мозга. Меняя активность γ-мотонейронов, мозговые центры влияют на чувствительность рецепторов нервно-мышечных веретён и, следовательно, на порог рефлекса растяжения. Сухожильные органы Гольджи. Обратный миотатический рефлекс Кроме нервно-мышечных веретён в активной части опорно-двигательного аппарата имеется ещё один тип проприрецепторов – сухожильные органы Гольджи. Сухожильные органы представляют собой сеть коллагеновых нитей, окружённую тонкой соединительнотканной капсулой. Диаметр сухожильного органа Гольджи составляет 100 мкм, длина – 1 мм. Коллагеновые нити внутри органа оплетённых нервными окончаниями, от которых отходят нервные волокна типа Ib. эти нервные волокна, как и Ia, соответствуют типу Aα по Гассеру-Эрлангеру. Сухожильные органы Гольджи расположены в месте прикрепления мышечных волокон к сухожильным, при этом каждый сухожильный орган связан с 3 – 25 (в среднем – с 10 – 15) мышечными волокнами (Рисунок 7). Таким образом, в отличии от нервно-мышечных веретён, которые ориентированы параллельно мышечным
волокнам, сухожильные органы по отношению к мышечным волокнам расположены последовательно.

Рисунок 6. Функция γ-петли
Следовательно, сухожильные органы реагируют не на изменение длины мышцы, а на развиваемое ей усилие, другими словами, на степень её напряжения. Афферентные нервные волокна от сухожильных органов Гольджи следуют в спинной мозг, где, в отличие от чувствительных волокон с нервно-мышечных веретён, переключаются на вставочные тормозные нейроны. Вставочные нейроны, в свою очередь, образуют синаптические контакты с α-мотонейронами гомонимной мышцы. Таким образом, активация сухожильных органов приводит к расслаблению мышцы, к которой крепится данное сухожилие. Описанный рефлекс называется обратным рефлексом растяжения (обратным миотатическим рефлексом) (Рисунок 8). Этот рефлекс, по-видимому, имеет две функции. Во-первых, он предупреждает чрезмерное напряжение скелетных мышц, которое может привести к тяжёлым травмам (разрыву мышцы, отрыву сухожилия от кости или от мышцы). Во-вторых, обратный миотатический рефлекс обеспечивает равномерность силы сокращения разных мышечных волокон в пределах одной мышцы. Те волокна, которые напряжены слишком сильно, расслабляются за счёт усиления тормозных влияний со стороны спинного мозга. Напряжение волокон, которые сокращаются недостаточно сильно, напротив, усиливается за счёт уменьшения тормозного воздействия.

Рисунок 7. Сухожильный орган Гольджи
Проприорецепторы посылают в головной мозг информацию о состоянии скелетно-мышечной системы Импульсация с проприорецепторов поступает не только в спинной мозг, но и по коллатералям афферентных волокон в различные отделы головного мозга. В частности, импульсы с нервно-мышечных веретён и сухожильных органов следуют по спинномозжечковым трактам в мозжечок, а также по системе задних столбов (тонкий и клиновидный пучок) в продолговатый мозг, затем – в таламус, а оттуда – в кору больших полушарий. Таким образом, мозжечок и кора большого мозга постоянно получают информацию о состоянии скелетно-мышечной системы, а именно о длине мышц и о степени их напряжения. Эта информация является необходимой как для регуляции произвольных движений и позы, так и для осознания положения частей тела в пространстве и друг относительно друга.

Рисунок 8. Обратный миотатический рефлекс с сухожильных органов Гольджи
Проприоцепция
Проприоцепция [ править | править код ]
Проприоцепция – это группа сигналов, посылаемых в центральную нервную систему специальными терминалами (проприоцепторами), расположенными в суставных капсулах, связках, сухожилиях и мышцах.
Проприоцепторы, среди которых выделяют мышечные рецепторы, или мышечные веретена (рецепторы растяжения), сухожильные рецепторы, или органы Гольджи (рецепторы мышечной силы), а также суставные рецепторы относятся к механорецепторам, посылающим в ЦНС информацию о положении, деформации и смещениях различных частей тела.
Функционирование этих рецепторов обеспечивает координацию всех подвижных органов и тканей животного и человека в состоянии покоя и во время любых двигательных актов. При экспериментальном выключении проприоцепторов животные теряют способность поддерживать естественные позы, двигаться и целесообразно реагировать на внешние воздействия.
Если человек закроет глаза и попытается написать текст, то буквы будут написаны достаточно четко. Этим простым способом легко убедиться в умении человека пользоваться информацией, идущей от мышц и суставов.
Проприоцепторы составляют периферическую часть проприоцептивной сенсорной системы, или двигательного анализатора. Вместе с тем, несмотря на то, что миллионы людей ежедневно пользуются услугами этого анализатора, мы до сегодняшнего дня знаем сравнительно мало о его деятельности. Это особенно касается работы коркового отдела двигательного анализатора. Внутренние проприоцепторы находятся в мышцах, сухожилиях, сухожильных влагалищах, межкостных мембранах, фасциях, тканях суставов, надкостнице и т.д. Среди них имеются неспециализированные рецепторы, встречающиеся и в других частях тела (свободные нервные окончания, инкапсулированные рецепторы типа телец Руффини и Пачини), и специализированные — мышечные веретена и сухожильные органы (или рецепторы) Гольджи.
Двигательный или кинестетический, анализатор (мышечная сенсорная система) обеспечивает формирование так называемого мышечного чувства при изменении напряжения мышц, суставных сумок, связок и сухожилий. Проблема мышечно-суставных ощущений имеет исключительное значение для физиологии и психологии. Специфические особенности человека появились благодаря более совершенной организации мышечной сенсорной системы по сравнению с животными.
И.М. Сеченов считал, что мышечное чувство является ближайшим регулятором движений и одним из орудий ориентации человека в пространстве и времени. Ощущения положения и перемещения тела в пространстве, ощущения во время трудовой деятельности и членораздельная речь лежат в основе формирования сознания человека и его представлений об окружающем мире.
Мышечное чувство обладает тремя качествами. Это, во-первых, ощущение положения конечностей, когда человек может определить положение своих конечностей и их частей относительно друг друга. Во-вторых, ощущение движения, когда, изменяя угол сгибания в суставе, человек осознаёт скорость и направление движения. Третьим качеством является ощущение усилия, когда человек может оценить мышечную силу, нужную для движения или удерживания суставов в определённом положении при подъёме или перемещении груза.
Наряду с кожной, зрительной и вестибулярной сенсорными системами двигательный анализатор оценивает положение тела в пространстве, позу, участвует в координации мышечной деятельности.
Двигательная сенсорная система служит для анализа состояния двигательного аппарата — его движения и положения. Информация о степени сокращения скелетных мышц, натяжении сухожилий, изменении суставных углов необходима для регуляции двигательных актов и поз.
Общий план организации. Двигательная сенсорная система состоит из следующих 3-х отделов:
К проприорецепторам относятся мышечные веретена, сухожильные органы (или органы Гольджи) и суставные рецепторы (рецепторы суставной капсулы и суставных связок). Все эти рецепторы представляют собой механорецепторы, специфическим раздражителем которых является их растяжение.
Мышечные веретена человека, представляют собой продолговатые образования длиной несколько миллиметров, шириной десятые доли миллиметра, которые расположены в толще мышцы. В разных скелетных мышцах число веретен на1 г ткани варьирует от нескольких единиц до сотни.
Таким образом, мышечные веретена как датчики состояния силы мышцы и скорости ее растяжения реагируют на два воздействия: периферическое — изменение длины мышцы, и центральное — изменение уровня активации гамма-мотонейронов. Поэтому реакции веретен в условиях естественной деятельности мышц довольно сложны. При растяжении пассивной мышцы наблюдается активация рецепторов веретен; она вызывает миотатичес-кий рефлекс, или рефлекс на растяжение. При активном сокращении мышцы уменьшение ее длины оказывает на рецепторы веретена дезактивирующее действие, а возбуждение гамма-мотонейронов, сопутствующее возбуждению альфа-мотонейронов, приводит к реактивации рецепторов. Вследствие этого импульсация от рецепторов веретен во время движения зависит от длины мышцы, скорости ее укорочения и силы сокращения.
Сухожильные органы (рецепторы) Гольджи человека располагаются в зоне соединения мышечных волокон с сухожилием, последовательно по отношению к мышечным волокнам.
Сухожильные органы представляют собой структуру вытянутой веретенообразной или цилиндрической формы, длина которой у человека может достигать1 мм. Этот первичночувствующий рецептор. В условиях покоя, т.е. когда мышца не сокращена, от сухожильного органа идет фоновая импульсация. В условиях мышечного сокращения частота импульсации возрастает прямо пропорционально величине мышечного сокращения, что позволяет рассматривать сухожильный орган как источник информации о силе, развиваемой мышцей. В тоже время сухожильный орган слабо реагирует на растяжение мышцы.
В результате последовательного крепления сухожильных органов к мышечным волокнам (а в ряде случаев — к мышечным веретенам), растяжение сухожильных механорецепторов происходит при напряжении мышц. Таким образом, в отличие от мышечных веретен, сухожильные рецепторы информируют нервные центры о степени напряжения мыши, и скорости его развития.
Эфферентная иннервация скелетной мышцы Эфферентная иннервация скелетной мышцы: 1 — сухожилие мышцы; 2 — сухожильные рецепторы Гольджи; 3 — свободные окончания; 4 — тельца Фатера-Пачини; 5 — афферентные волокна; 6 — эфферентные волокна; 7 — мышечное веретено; 8 — окончания афферентных волокон
Суставные рецепторы реагируют на положение сустава и на изменения суставного угла, участвуя, таким образом, в системе обратных связей от двигательного аппарата и в управлении им. Суставные рецепторы информируют о положении отдельных частей тела в пространстве и относительно друг друга. Эти рецепторы представляют собой свободные нервные окончания или окончания, заключенные в специальную капсулу. Одни суставные рецепторы посылают информацию о величине суставного угла, т. е. о положении сустава. Их импульсация продолжается в течение всего периода сохранения данного угла. Она тем большей частоты, чем больше сдвиг угла. Другие суставные рецепторы возбуждаются только в момент движения в суставе, т. е. посылают информацию о скорости движения. Частота их импульсации возрастаете увеличением скорости изменения суставного угла.
Проводниковый и корковый отделы проприоцеп-тивного анализатора млекопитающих и человека. Информация от мышечных, сухожильных и суставных рецепторов поступает через аксоны первых афферентных нейронов, находящихся в спинномозговых ганглиях, в спинной мозг, где частично переключается на альфа-мотонейроны или на вставочные нейроны (например, на клетки Реншоу), а частично направляется по восходящим путям в высшие отделы головного мозга. В частности, по путям Флексига и Говерса проприоцептивная импульсация доставляется к мозжечку, а по пучкам Голля и Бурдаха, проходящим в дорсальных канатиках спинного мозга, она доходит до нейронов одноименных ядер, расположенных в продолговатом мозге.
Аксоны таламических нейронов (нейронов третьего порядка) оканчиваются в коре больших полушарий, главным образом, в соматосенсорной коре (постцентральная извилина) и в области сильвие-вой борозды (соответственно, участки S-1 и S-2), а также частично в двигательной (префронтальной) области коры. Эта информация используется двигательными системами мозга достаточно широко, в том числе для принятия решения о замысле движения, а также для его реализации. Кроме того, у человека на основе про-приоцептивной информации формируются представления о состоянии мышц и суставов, а также, в целом, о положении тела в пространстве.
Сигналы, идущие от рецепторов мышечных веретен, сухожильных органов, суставных сумок и тактильных рецепторов кожи, называют кинестетическими, т. е. информирующими о движении тела. Их участие в произвольной регуляции движений различно. Сигналы от суставных рецепторов вызывают заметную реакцию в коре больших полушарий и хорошо осознаются. Благодаря им человек лучше воспринимает различия при движениях в суставах, чем различия в степени напряжения мышц при статических положениях или поддержании веса. Сигналы же от других проприорецепторов, поступающие преимущественно в мозжечок, обеспечивают бессознательную регуляцию, подсознательный контроль движений и поз.
Таким, образом, проприоцептивные ощущения дают человеку возможность воспринимать изменения положения отдельных частей тела в покое и во время совершаемых движений. Информация, поступающая от проприоцепторов, позволяет ему постоянно контролировать позу и точность произвольных движений, дозировать силу мышечных сокращений при противодействии внешнему сопротивлению, например при подъеме или перемещении груза.
Проприоцептивное восприятие положения тела и движений происходит в результате объединения в соматосенсорной коре информации от всех разновидностей проприоцепторов.
Вестибулярный анализатор – анализатор, обеспечивающий анализ информации о положении и перемещениях тела в пространстве.



