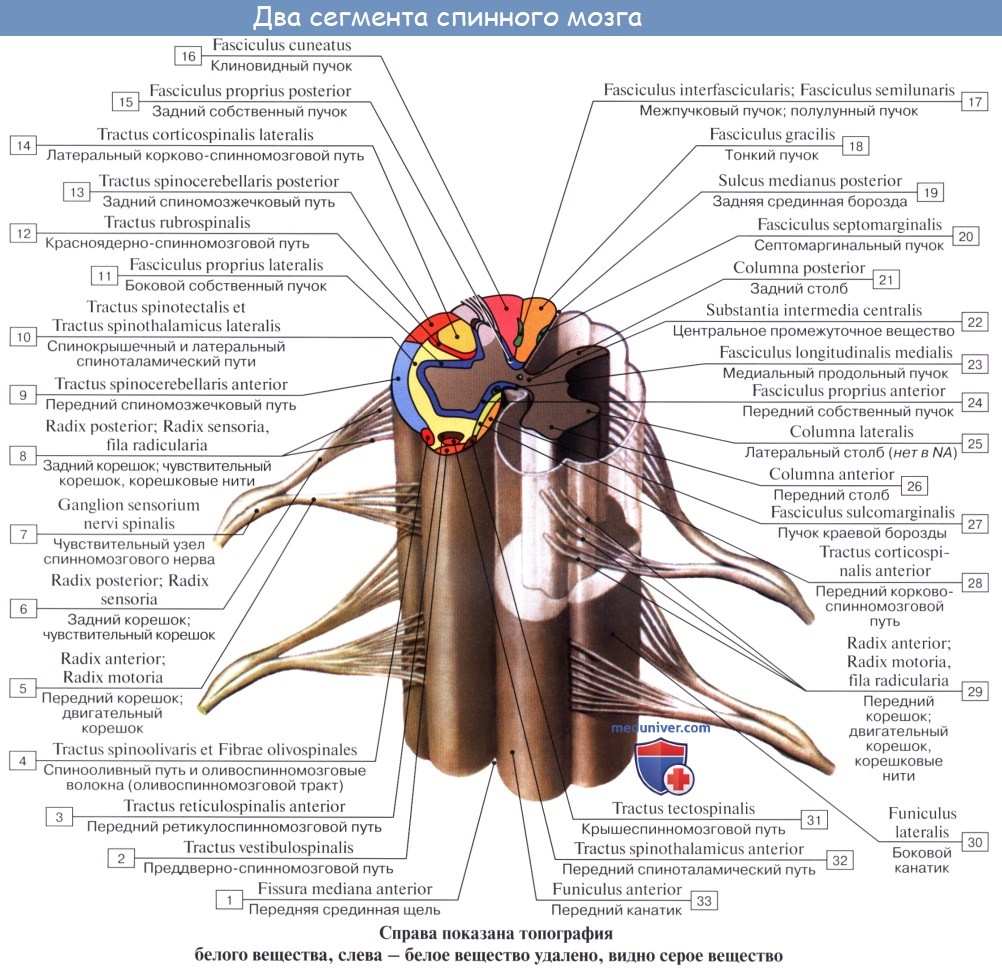
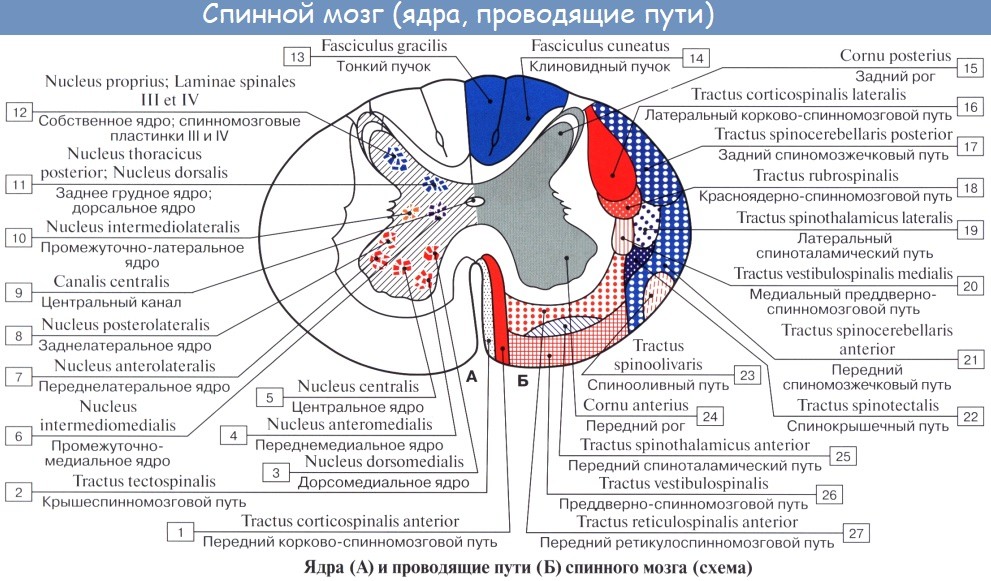
От коры головного мозга: 1) передний корково-спинномозговой (пирамидный) путь, tractus corticospinalis (pyramidalis) anterior, составляет с латеральным пирамидным пучком общую пирамидную систему.
От среднего мозга: 2) tractus tectospinal, лежит медиальнее пирамидного пучка, ограничивая fissura mediana anterior. Благодаря ему осуществляются рефлекторные защитные движения при зрительных и слуховых раздражениях — зрительно-слуховой рефлекторный тракт.
Ряд пучков идет к передним рогам спинного мозга от различных ядер продолговатого мозга, имеющих отношение к равновесию и координации движений, а именно:
3) от ядер вестибулярного нерва — tractus vestibulospinal — лежит на границе переднего и бокового канатиков;
4) от formatio reticularis — tractus reticulospindlis anterior, лежит в средней части переднего канатика;
5) собственно пучки, fasciculi proprii, непосредственно прилегают к серому веществу и относятся к собственному аппарату спинного мозга.
Эти борозды делят каждую половину белого вещества спинного мозга на три продольных канатика: передний — funiculus anterior, боковой— funiculus lateralis и задний — funiculus posterior. Задний канатик в шейном и верхнегрудном отделах делится еще промежуточной бороздкой, sulcus intermedius posterior, на два пучка: fasciculus gracilis и fasciculus cuneatus. Оба эти пучка под теми же названиями переходят вверху на заднюю сторону продолговатого мозга.
На той и другой стороне из спинного мозга выходят двумя продольными рядами корешки спинномозговых нервов. Передний корешок, radix ventral is s. anterior, выходящий через sulcus anterolateralis, состоит из нейритов двигательных (центробежных, или эфферентных) нейронов, клеточные тела которых лежат в спинном мозге, тогда как задний корешок, radix dorsalis s. posterior, входящий в sulcus posterolateralis, содержит отростки чувствительных (центростремительных, или афферентных) нейронов, тела которых лежат в спинномозговых узлах.
В межпозвоночных отверстиях вблизи места соединения обоих корешков задний корешок имеет утолщение — спинномозговой узел, ganglion spinale, содержащий ложноуниполярные нервные клетки (афферентные нейроны) с одним отростком, который делится затем на две ветви: одна из них, центральная, идет в составе заднего корешка в спинной мозг, другая, периферическая, продолжается в спинномозговой нерв. Таким образом, в спинномозговых узлах отсутствуют синапсы, так как здесь лежат клеточные тела только афферентных нейронов. Этим названные узлы отличаются от вегетативных узлов периферической нервной системы, так как в последних вступают в контакты вставочные и эфферентные нейроны. Спинномозговые узлы крестцовых корешков лежат внутри крестцового канала, а узел копчикового корешка — внутри мешка твердой оболочки спинного мозга.
Вследствие того что спинной мозг короче позвоночного канала, место выхода нервных корешков не соответствует уровню межпозвоночных отверстий. Чтобы попасть в последние, корешки направляются не только в стороны от мозга, но еще и вниз, при этом тем отвеснее, чем ниже они отходят от спинного мозга. В поясничной части последнего нервные корешки спускаются к соответствующим межпозвоночным отверстиям параллельно filum terminate, облекая ее и conus medullaris густым пучком, который носит название конского хвоста, cauda equina.
Задние канатики содержат волокна задних корешков спинномозговых нервов, слагающиеся в две системы:
1. Медиально расположенный тонкий пучок, fasciculus gracilis.
2. Латерально расположенный клиновидный пучок, fasciculus cuneatus. Пучки тонкий и клиновидный проводят от соответствующих частей тела к коре головного мозга сознательную проприоцептивную (мышечно-суставное чувство) и кожную (чувство стереогноза — узнавание предметов на ощупь) чувствительность, имеющую отношение к определению положения тела в пространстве, а также тактильную чувствительность.
Боковые канатики содержат следующие пучки:
К заднему мозгу: 1) tractus spinocerebellaris posterior, задний спинно-мозжечковый путь, располагается в задней части бокового канатика по его периферии; 2) tractus spinocerebellaris anterior, передний спинномозжечковый путь, лежит вентральнее предыдущего.
Оба спинно-мозжечковых тракта проводят бессознательные проприоцептивные импульсы (бессознательная координация движений).
К среднему мозгу: 3) tractus spinotectalis, спинно-покрышечный путь, прилегает к медиальной стороне и передней части tractus spinocerebellaris anterior.
К промежуточному мозгу: 4) tractus spinothalamicus lateralis прилегает с медиальной стороны к tractus spinocerebellaris anterior, тотчас позади tractus spinotectalis. Он проводит в дорсальной части тракта тем-‘ пературные раздражения, а в вентральной — болевые; 5) tractus spinothalamicus anteriror s. ventralis аналогичен предыдущему, но располагается кпереди от соименного латерального и является путем проведения импульсов осязания, прикосновения (тактильная чувствительность). По последним данным, этот тракт располагается в переднем канатике.
От коры большого мозга: 1) латеральный корково-спинномозговой (пирамидный) путь, tractus corticospinalis (pyramidalis) lateralis. Этот тракт является сознательным эфферентным двигательным путем.
От среднего мозга: 2) tractus rubrospinal. Он является бессознательным эфферентным двигательным путем.
От заднего мозга: 3) tractus olivospinal, лежит вентральнее tractus spinocerebellaris anterior, вблизи переднего канатика.
Белое вещество, substantia alba,спинного мозга состоит из нервных отростков, которые составляют три системы нервных волокон:
1. Короткие пучки ассоциативных волокон, соединяющих участки спинного мозга на различных уровнях (афферентные и вставочные нейроны). 2. Длинные центростремительные (чувствительные, афферентные). 3. Длинные центробежные (двигательные, эфферентные).
Первая система (коротких волокон) относится к собственному аппарату спинного мозга, а остальные две (длинных волокон) составляют проводниковый аппарат двусторонних связей с головным мозгом.
Собственный аппарат включает серое вещество спинного мозга с задними и передними корешками и собственными пучками белого вещества (fasciculi proprii), окаймляющими серое в виде узкой полосы. По развитию собственный аппарат является образованием филогенетически более старым и потому сохраняет примитивные черты строения — сегментарность, отчего его называют также сегментарным аппаратом спинного мозга в отличие от остального несегментированного аппарата двусторонних связей с головным мозгом.
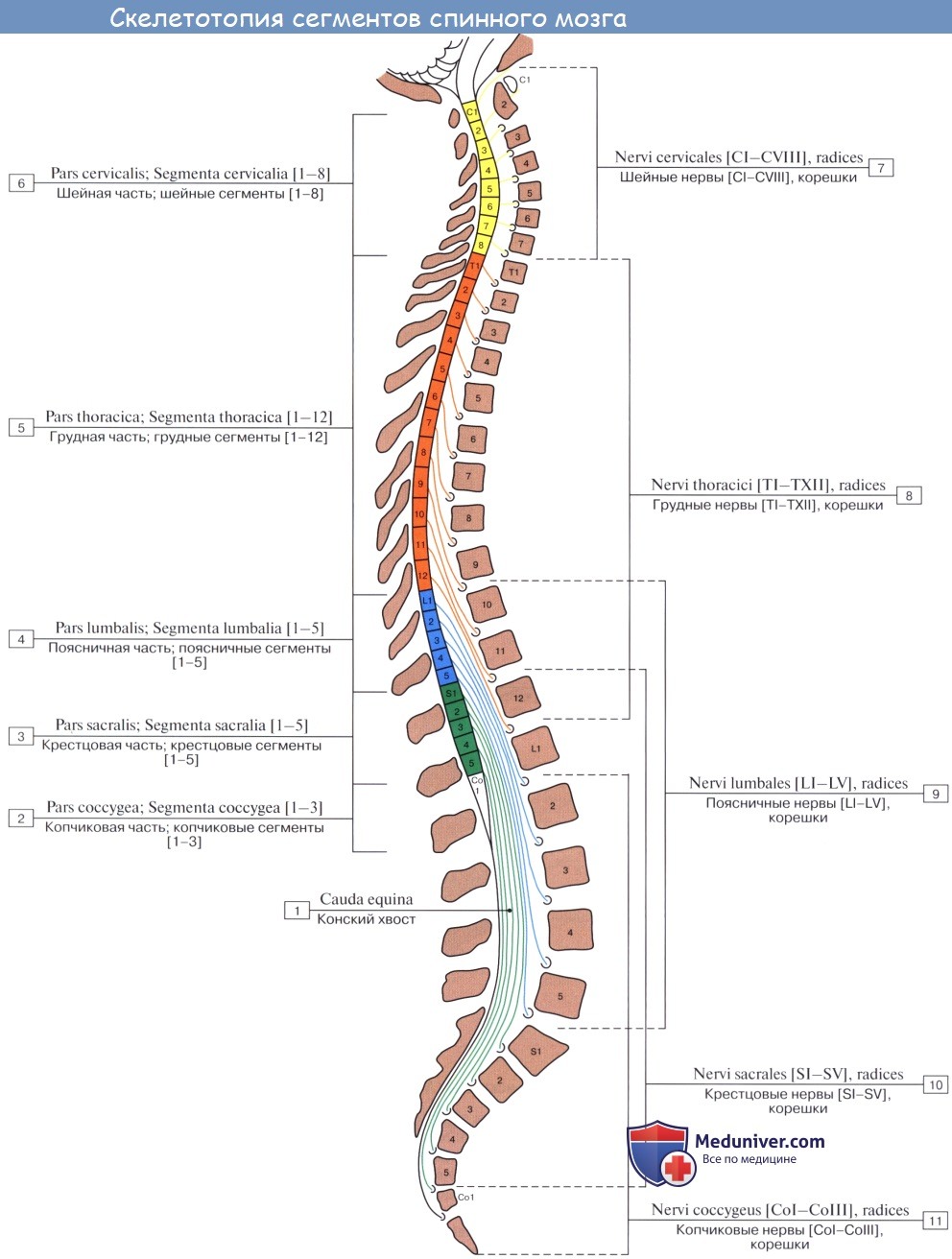
Таким образом, нервный сегмент — это поперечный отрезок спинного мозга и связанных с ним правого и левого спинномозговых нервов, развившихся из одного невротома (невромера). Он состоит из горизонтального слоя белого и серого вещества (задние, передние и боковые рога), содержащего нейроны, отростки которых проходят в одном парном (правом и левом) спинномозговом нерве и его корешках. В спинном мозге различают 31 сегмент, которые топографически делятся на 8 шейных, 12 грудных, 5 поясничных, 5 крестцовых и 1 копчиковый. В пределах нервного сегмента замыкается короткая рефлекторная дуга.
Так как собственный сегментарный аппарат спинного мозга возник тогда, когда еще не было головного, то функция его — это осуществление тех реакций в ответ на внешнее и внутреннее раздражения, которые в процессе эволюции возникли раньше, т. е. врожденных реакций.
Аппарат двусторонних связей с головным мозгом филогенетически более молодой, так как возник лишь тогда, когда появился головной мозг.
По мере развития последнего разрастались кнаружи и проводящие пути, связывающие спинной мозг с головным. Этим объясняется тот факт, что белое вещество спинного мозга как бы окружило со всех сторон серое вещество. Благодаря проводниковому аппарату собственный аппарат спинного мозга связан с аппаратом головного мозга, который объединяет работу всей нервной системы. Нервные волокна группируются в пучки, а из пучков составляются видимые невооруженным глазом канатики: задний, боковой и передний. В заднем канатике, прилежащем к заднему (чувствительному) рогу, лежат пучки восходящих нервных волокон; в переднем канатике, прилежащем к переднему (двигательному) рогу, лежат пучки нисходящих нервных волокон; наконец, в боковом канатике находятся и те и другие. Кроме канатиков, белое вещество находится в белой спайке, comissura alba, образующейся вследствие перекреста волокон спереди от substantia intermedia centralis; сзади белая спайка отсутствует.
В переднем канатике спинного мозга расположен
Существуют следующие нисходящие проводящие пути: • корково-спинномозговой проводящий путь (пирамидный проводящий путь); • ретикуло-спинномозговой проводящий путь (экстрапира-мидный путь); • преддверно-спинномозговой проводящий путь; • покрышечно-спинномозговой проводящий путь; • шовно-спинномозговой проводящий путь; • проводящие пути аминергических систем ЦНС; • проводящие пути вегетативной нервной системы.
Корково-спинномозговой проводящий путь
Корково-спинномозговой проводящий путь представляет собой крупный проводящий путь произвольной двигательной активности. Около 40 % его волокон начинается из первичной моторной коры прецентральной извилины. Остальные волокна берут начало из дополнительной моторной области на медиальной стороне полушария, премоторной коры головного мозга на латеральной стороне полушария, соматической сенсорной коры, коры теменной доли и коры поясной извилины. Волокна от двух вышеупомянутых сенсорных центров заканчиваются на чувствительных ядрах ствола головного мозга и спинного мозга, где они регулируют передачу чувствительных импульсов.
Корково-спинномозговой проводящий путь спускается вниз через лучистый венец и заднюю ножку внутренней капсулы к стволу головного мозга. Затем он проходит в ножке (головного мозга) на уровне среднего мозга и базилярной части моста, достигая продолговатого мозга. Здесь он образует пирамиду (отсюда название — пирамидный проводящий путь).
Демонстрация хода волокон пирамидного пути с левой стороны. Дополнительная моторная область на медиальной стороне полушария. Стрелкой показан уровень перекреста пирамид. Чувствительные нейроны выделены синим цветом.Коронарный срез бальзамированного головного мозга пациента с последующей обработкой сульфатом меди (окраска по Маллигану), демонстрирующий неокрашенные корково-спинномозговые волокна, идущие через ядра моста в сторону пирамид.
Характеристика волокон корково-спинномозгового пути выше уровня спинномозгового перехода:
• около 80 % (70-90 %) волокон переходят на противоположную сторону на уровне перекреста пирамид;
• эти волокна спускаются по противоположной стороне спинного мозга и составляют латеральный корково-спинномозговой проводящий путь (перекрещивающийся корково-спинномозговой проводящий путь); оставшиеся 20 % волокон не перекрещиваются и продолжают спускаться вниз в передней части спинного мозга;
• половина из этих неперекрещивающихся волокон вступает в передний/вентральный корково-спинномозговой проводящий путь и располагается в вентральном/переднем канатике спинного мозга на шейном и верхнем грудном уровнях; данные волокна переходят на противоположную сторону на уровне белой спайки и иннервируют мышцы передней и задней стенок брюшной полости;
• другая половина вступает в латеральный корково-спинномозговой проводящий путь на своей половине спинного мозга.
Считают, что корково-спинномозговой проводящий путь содержит около 1 млн. нервных волокон. Средняя скорость проведения импульса составляет 60 м/с, что указывает на средний диаметр волокна, равный 10 мкм («правило шести»). Около 3 % волокон — очень крупные (до 20 мкм); они отходят от гигантских нейронов (клетки Беца), расположенных в основном в области двигательной коры, отвечающей за иннервацию нижних конечностей. Все волокна корково-спинномозгового пути — возбуждающие и в качестве медиатора используют глутамат.
а) Мотонейроны дистальных отделов конечностей. В передних рогах серого вещества спинного мозга аксоны латерального корково-спинномозгового пути могут непосредственно образовывать синапсы на дендритах α- и γ-мотонейронов, иннервирующих мышцы конечностей, особенно верхних (однако, как правило, это происходит через интернейроны в пределах серого вещества спинного мозга). Отдельные аксоны латерального корково-спинномозгового пути могут активировать «большие» или «малые» двигательные единицы.
Двигательная единица — это комплекс, состоящий из нейрона переднего рога спинного мозга и всех мышечных волокон, которые этот нейрон иннервирует. Нейроны малых двигательных единиц избирательно иннервируют небольшое количество мышечных волокон и участвуют в выполнении тонких и точных движений (например, при игре на пианино). Нейроны переднего рога, иннервирующие крупные мышцы (например, большую ягодичную мышцу), способны по отдельности вызвать сокращение сотни мышечных клеток сразу, так эти мышцы отвечают за грубые и простые движения.
Уникальное свойство этих корковомотонейронных волокон латерального корково-спинномозгового пути демонстрирует понятие «фракционирования», относящееся к переменной активности интернейронов, в результате чего небольшие группы нейронов могут быть избирательно активированы для выполнения конкретной общей функции. Это легко показать на указательном пальце, который может быть согнут или разогнут независимо от положения других пальцев (хотя три из его длинных сухожилий имеют общее начало с мышечным ложем всех четырех пальцев).
Фракционирование имеет большое значение при выполнении привычных движений, таких как застегивание пальто или завязывание шнурков. Травматическое или другое повреждение корковомотонейронной системы на любом уровне влечет за собой утрату навыков выполнения привычных движений, которые затем редко поддаются восстановлению.
При выполнении данных движений α- и γ-мотонейроны активируются совместно через латеральный корково-спинномозговой проводящий путь таким образом, что веретена мышц, первично задействованных в движении, посылают импульсы об активном растяжении, а веретена мышц-антагонистов — о пассивном растяжении.
Продолговатый мозг и верхние отделы спинного мозга, вид спереди. Продемонстрированы три группы нервных волокон левой пирамиды.
б) Клетки Реншоу. Функции синапсов латерального корково-спинномозгового пути на клетках Реншоу довольно многочисленны, так как торможение на некоторых клеточных синапсах главным образом происходит за счет интернейронов типа Iа; на других синапсах данную функцию выполняют клетки Реншоу. Вероятно, наиболее важная функция — контроль совместного сокращения основных движущих мышц и их антагонистов для фиксации одного или нескольких суставов, например при работе с кухонным ножом или лопатой. Совместное сокращение происходит за счет инактивации ингибирующих интернейронов Iа клетками Реншоу.
в) Возбуждающие интернейроны. Латеральный корково-спинно-мозговой проводящий путь влияет на деятельность двигательных нейронов, расположенных в средней части серого вещества и в основании переднего рога спинного мозга, иннервирующих осевые (позвоночные) мышцы и мышцы проксимальных отделов конечностей посредством возбуждающих интернейронов. г) la-ингибирующие интернейроны. Эти нейроны также расположены в средней части серого вещества спинного мозга и активируются латеральным корково-спинномозговым путем в первую очередь при совершении произвольных движений.
Активность Ia-интернейронов способствует расслаблению мышц-антагонистов до того, как начнут сокращаться мышцы-агонисты. Кроме того, они вызывают рефрактерность мотонейронов мышц-антагонистов к стимуляции афферентами нервно-мышечного веретена при их пассивном растяжении во время движения. Последовательность процессов при произвольном сгибания коленного сустава показана на рисунке ниже.
(Обратите внимание на терминологию: в спокойном положении стоя колени человека «закрыты» в небольшом переразгибании, а четырехглавая мышца бедра находится в неактивном состоянии, о чем свидетельствует «свободное» положение надколенника. При попытке сгибания одного или обоих колен происходит подергивание четырехглавой мышцы бедра в ответ на пассивное растяжение в ней десятков мышечных веретен. Поскольку таким образом происходит сопротивление сгибанию, рефлекс называют рефлексом сопротивления.
С другой стороны, во время произвольного сгибания коленного сустава мышцы способствуют данному движению с помощью такого же механизма, но уже через рефлекс помощи. Изменение знака с отрицательного на положительный называют рефлексом перемены направления.)
д) Пресинаптические ингибиторные нейроны, обеспечивающие рефлекс растяжения. Рассмотрим движения спринтера. На каждом шаге сила тяжести тянет его тело вниз, на выпрямленное четырехглавой мышцей колено. В момент соприкосновения с землей все нервно-мышечные веретена в сокращенной четырехглавой мышце резко растягиваются, в результате чего возникает опасность разрыва мышцы. Сухожильный орган Гольджи обеспечивает некоторую защиту посредством внутреннего торможения, однако основной защитный механизм обеспечивает латеральный корково-спинномозговой путь через пресинаптическое торможение афферентов веретен вблизи их контакта с мотонейронами.
В то же время удлинение паузы до ахиллового рефлекса служит преимуществом в этой ситуации, так как происходит восстановление мотонейронов, иннервирующих заднюю часть голени, для следующего рывка. Предполагают, что степень подавления рефлекса растяжения со стороны латерального корково-спинномозгового пути зависит от конкретных движений.
е) Пресинаптическое ингибирование чувствительных нейронов первого порядка. В заднем роге серого вещества спинного мозга существует некоторое подавление передачи чувствительных импульсов в спиноталамический проводящий путь при совершении произвольных движений. Это происходит путем активации синапсов, образованных ингибирующими вставочными нейронами и первичными чувствительными нервными окончаниями.
Еще более тонкую регуляцию наблюдают на уровне тонкого и клиновидного ядер, где волокна пирамидного пути (после пересечения) способны усиливать передачу чувствительных импульсов во время медленных аккуратных движений или ослаблять ее во время совершения быстрых движений.
Последовательность событий при выполнении произвольного движения (сгибания колена). МН — мотонейроны. (1) Активация la интернейронов ингибирует их антагонисты-α-мотонейроны. (2) Активация агонистов α- и γ-мотонейронов. (3) Активация экстрафузальных и интрафузальных мышечных волокон. (4) Импульсация от активно растянутых нервно-мышечных веретен увеличивает активность агониста а-мотонейрона и снижает активность его антагонистов. (5) Iа-волокна от пассивно растянутых нервно-мышечных веретен-антагонистов направляются к соответствующим рефрактерным а-мотонейронам. Обратите внимание: последовательность «γ-мотонейронон—Ia-волокно—α-мотонейрон» образует γ-петлю.
Редактор: Искандер Милевски. Дата публикации: 15.11.2018
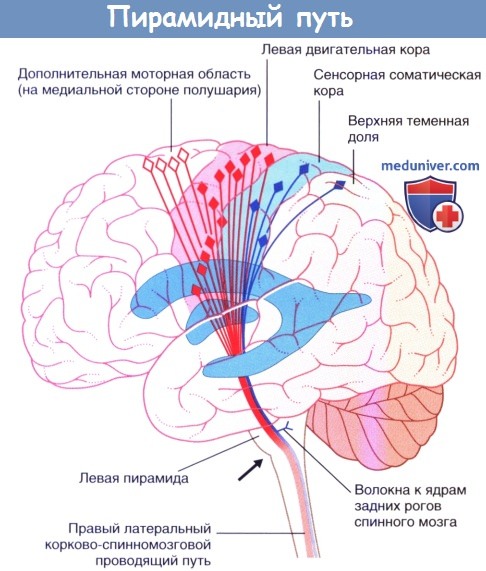 Демонстрация хода волокон пирамидного пути с левой стороны.
Демонстрация хода волокон пирамидного пути с левой стороны. 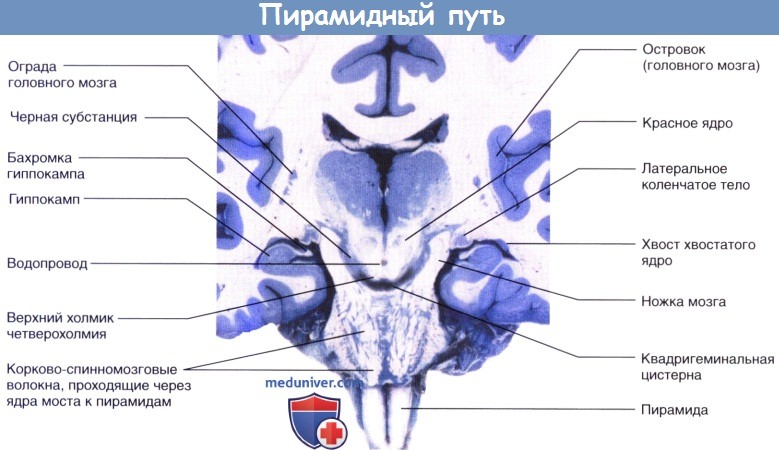 Коронарный срез бальзамированного головного мозга пациента с последующей обработкой сульфатом меди (окраска по Маллигану),
Коронарный срез бальзамированного головного мозга пациента с последующей обработкой сульфатом меди (окраска по Маллигану), 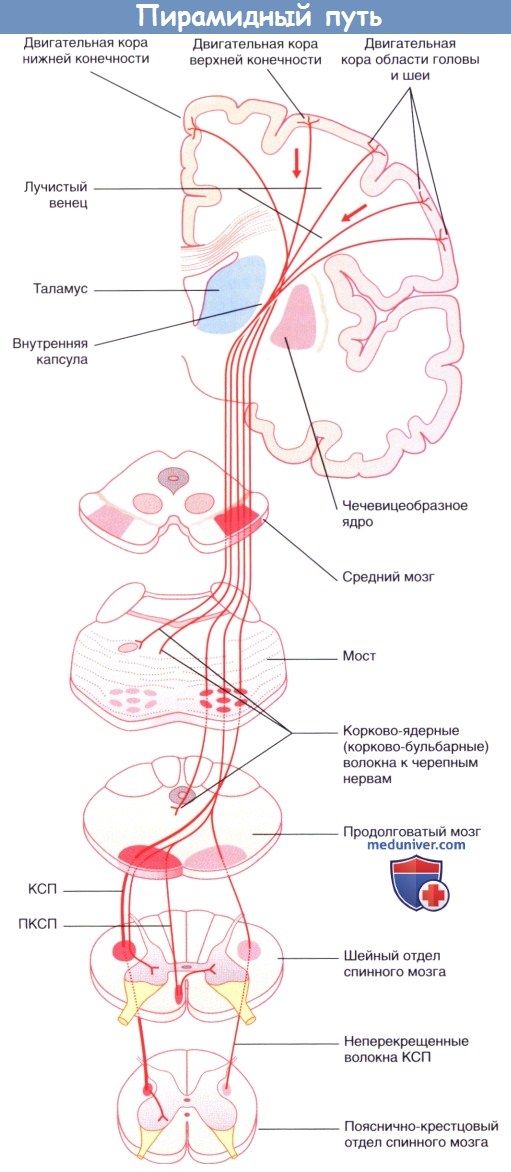 Пирамидный проводящий путь.
Пирамидный проводящий путь. 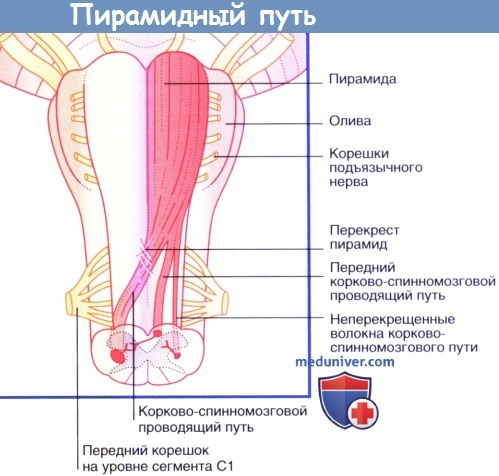 Продолговатый мозг и верхние отделы спинного мозга, вид спереди.
Продолговатый мозг и верхние отделы спинного мозга, вид спереди. 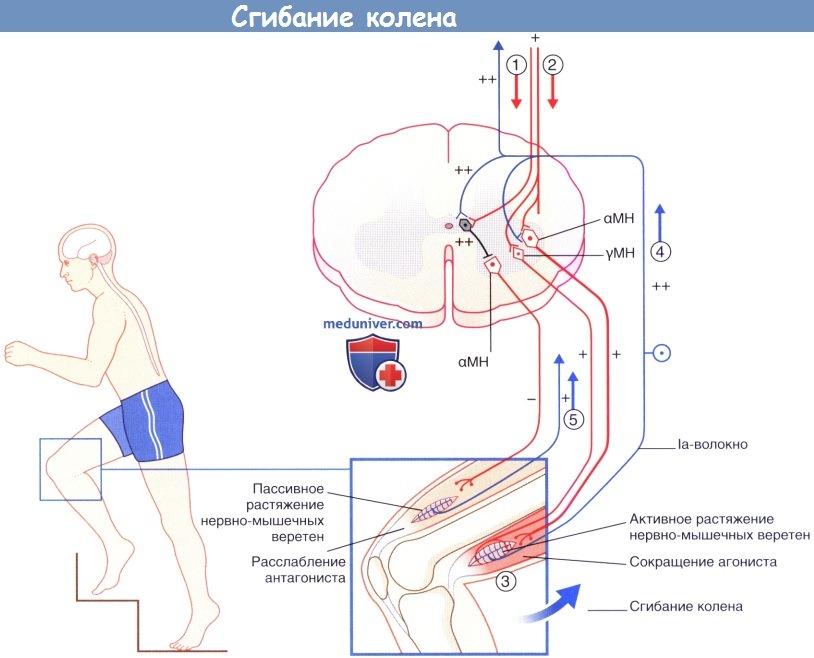 Последовательность событий при выполнении произвольного движения (сгибания колена). МН — мотонейроны.
Последовательность событий при выполнении произвольного движения (сгибания колена). МН — мотонейроны. 


