в промежуточном мозге выделяют таламус и гипоталамус
В промежуточном мозге выделяют таламус и гипоталамус
Рекомендуем:
Анатомия человека:
Анатомия человека
Анатомическая терминология
Анатомия костей и суставов
Анатомия мышц
Анатомия внутренних органов
Анатомия эндокринных органов
Анатомия сердца и сосудов
Анатомия нервной системы
Анатомия органов чувств
Видео по анатомии
Книги по анатомии
Топографическая анатомия
Форум
Оглавление темы «Передний мозг, prosencephalon. Промежуточный мозг, diencephalon.»:
Промежуточный мозг, diencephalon
1) дорсальную (филогенетически более молодую) — thalamencephalon — центр афферентных путей и
2) вентральную (филогенетически более старую) — hypothalamus — высший вегетативный центр. Полостью diencephalon является III желудочек.
Таламический мозг, thalamencephalon
Thalamencephalon в свою очередь состоит из трех частей: thalamus — таламус, epithalamus — надталамическая область и metathalamus — заталамическая область.
Видео №1: анатомия промежуточного мозга от А.А. Стрелкова
Редактор: Искандер Милевски. Дата последнего обновления публикации: 14.8.2020
В промежуточном мозге выделяют таламус и гипоталамус
Рекомендуем:
Анатомия человека:
Анатомия человека
Анатомическая терминология
Анатомия костей и суставов
Анатомия мышц
Анатомия внутренних органов
Анатомия эндокринных органов
Анатомия сердца и сосудов
Анатомия нервной системы
Анатомия органов чувств
Видео по анатомии
Книги по анатомии
Топографическая анатомия
Форум
Оглавление темы «Передний мозг, prosencephalon. Промежуточный мозг, diencephalon.»:
Таламус, thalamus. Строение таламуса. Ядра таламуса. Функции и значение таламуса
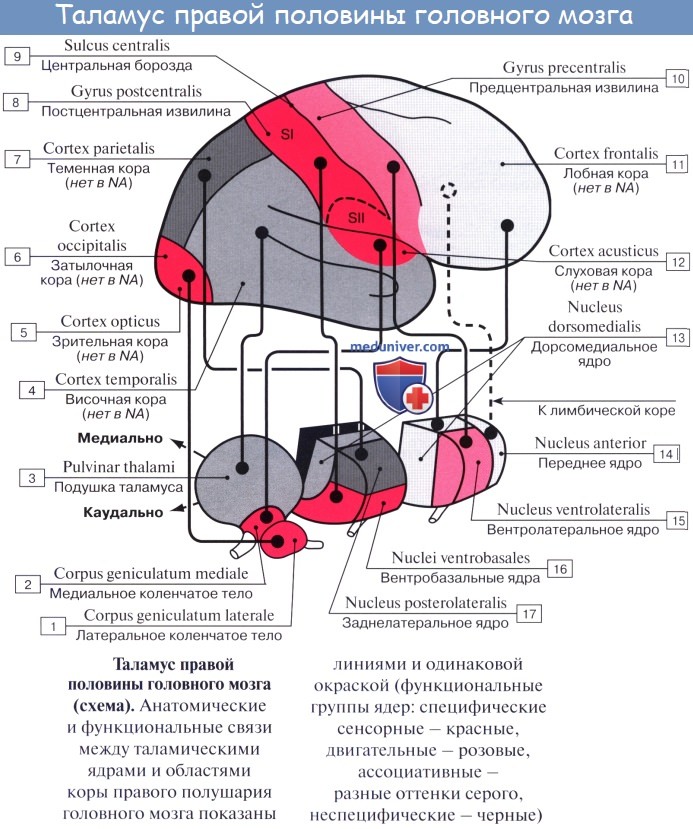
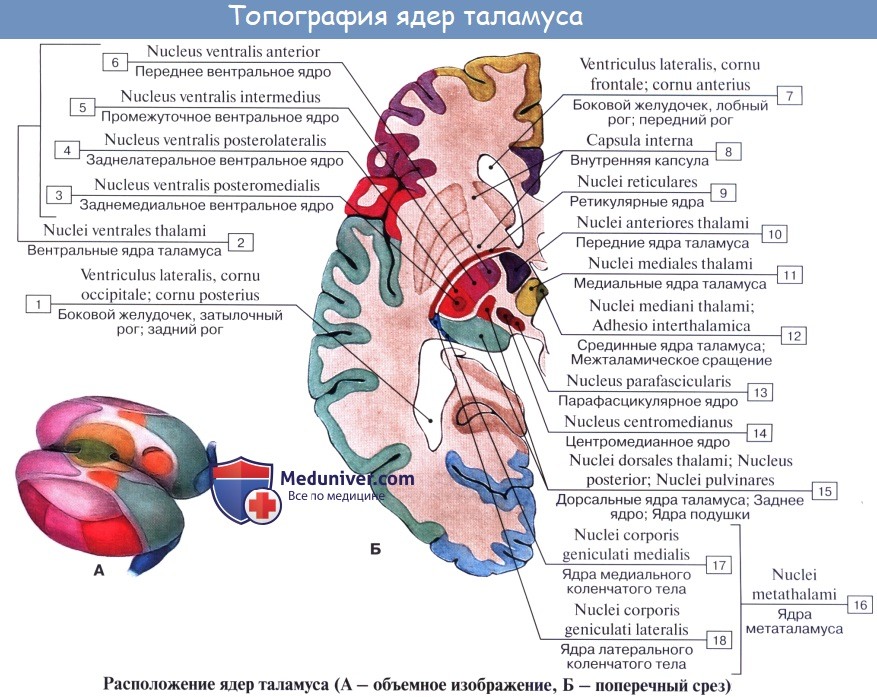
Thalamus, таламус, представляет собой большое парное скопление серого вещества в боковых стенках промежуточного мозга по бокам III желудочка, имеющее яйцевидную форму, причем передний его конец заострен в виде tuberculum anterius, а задний расширен и утолщен в виде подушки, pulvinar.
Деление на передний конец и подушку соответствует функциональному делению thalamus на центры афферентных путей (передний конец) и на зрительный центр (задний).
Дорсальная поверхность покрыта тонким слоем белого вещества — stratum zonule. В латеральном своем отделе она обращена в полость бокового желудочка, отделяясь от соседнего с ней хвостатого ядра пограничной бороздкой, sulcus terminalis, являющейся границей между telencephalon, к которому принадлежит хвостатое ядро, и diencephalon, к которому относится таламус. По этой бороздке проходит полоска мозгового вещества, stria terminalis.
Медиальная поверхность таламуса, покрытая тонким слоем серого вещества, расположена вертикально и обращена в полость III желудочка, образуя его латеральную стенку. Сверху она отграничивается от дорсальной поверхности посредством белой мозговой полоски, stria medullaris thalami. Обе медиальные поверхности таламусов соединены между собой серой спайкой — adhesio interthalamica, лежащей почти посередине. Латеральная поверхность таламуса граничит с внутренней капсулой, capsula interna.
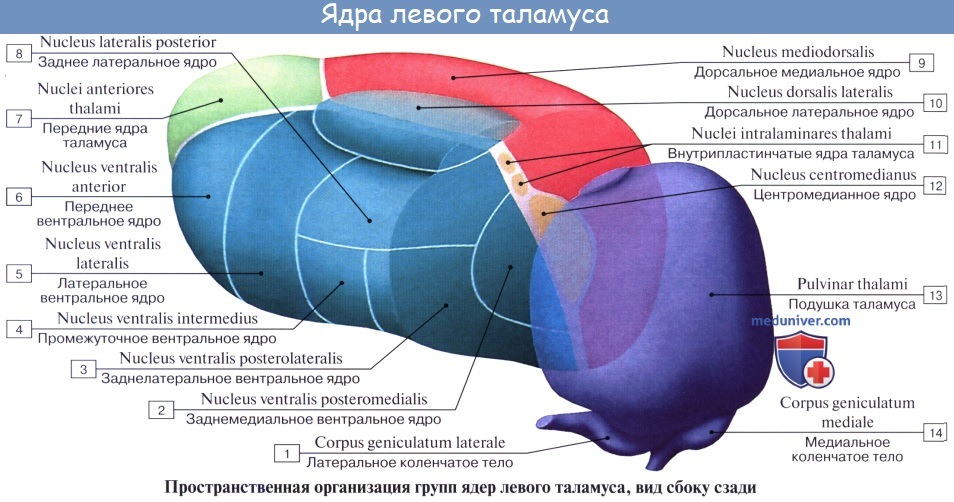
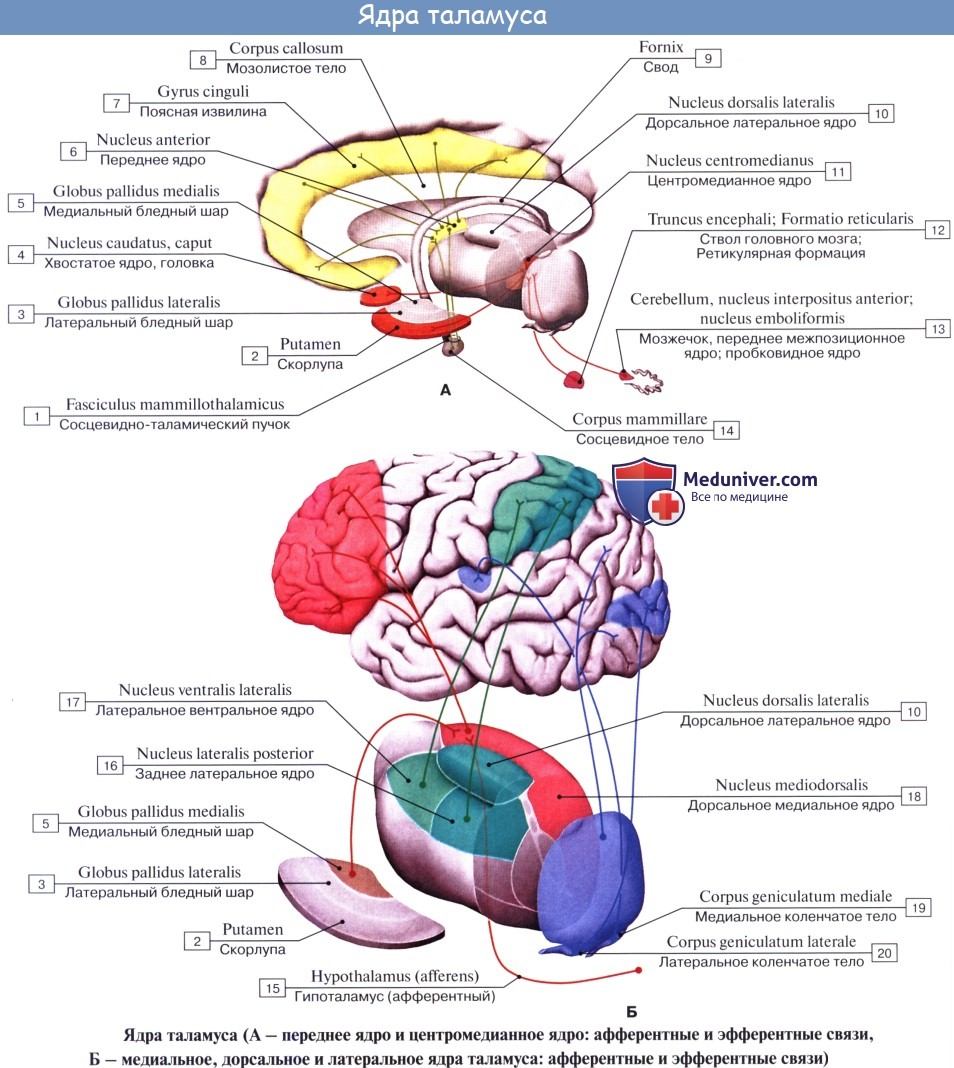
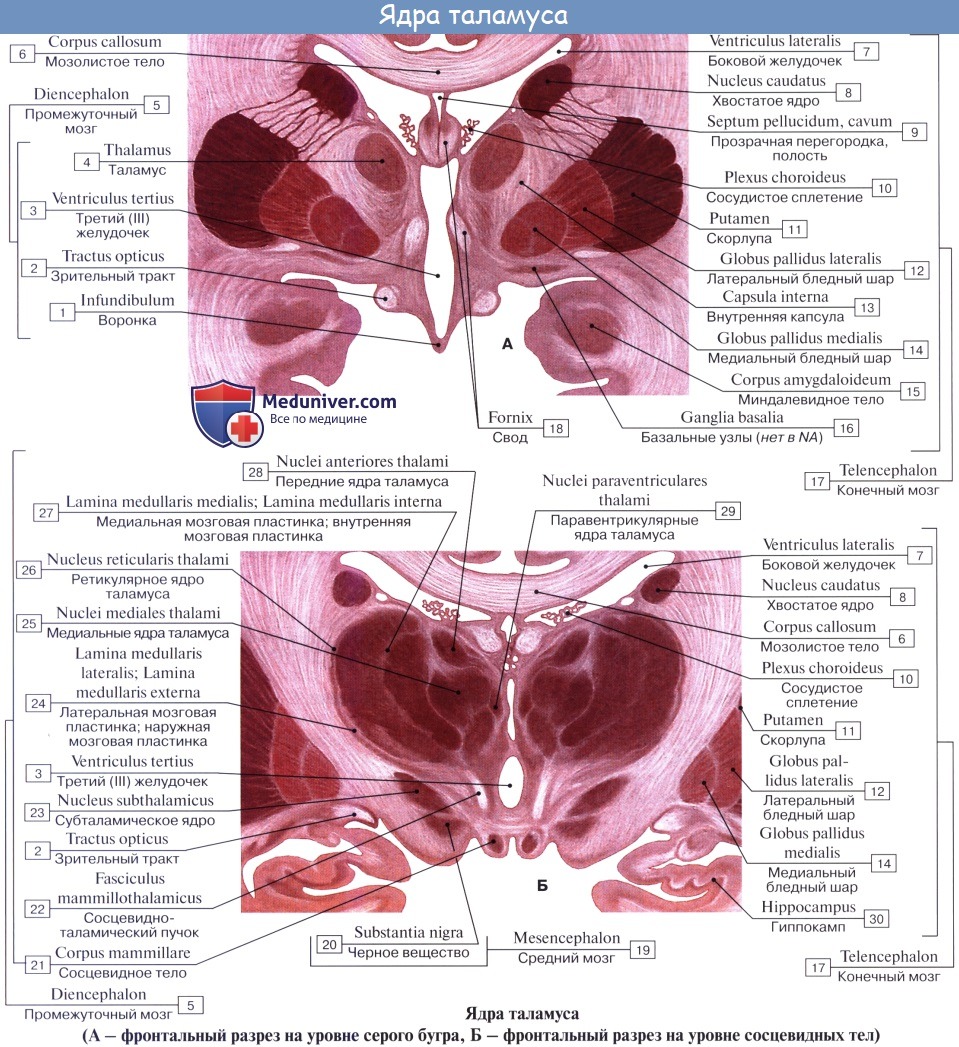
Нижней своей поверхностью таламус располагается над ножкой мозга, срастаясь с ее покрышкой. Как видно на разрезах, серая масса таламуса белыми прослойками, laminae medullares thalami, разделяется на отдельные ядра, носящие названия в зависимости от их топографии: передние, центральные, медиальные, латеральные, вентральные и задние.
Функциональное значение таламуса очень велико. В нем переключаются афферентные пути: в его подушке, pulvinar, где находится заднее ядро, оканчивается часть волокон зрительного тракта (подкорковый центр зрения, ассоциативное ядро таламуса), в передних ядрах — пучок, идущий от corpora mamillaria и связывающий таламус с обонятельной сферой, и, наконец, все остальные афферентные чувствительные пути от нижележащих отделов центральной нервной системы в остальных его ядрах, причем lemniscus medialis заканчивается в латеральных ядрах.
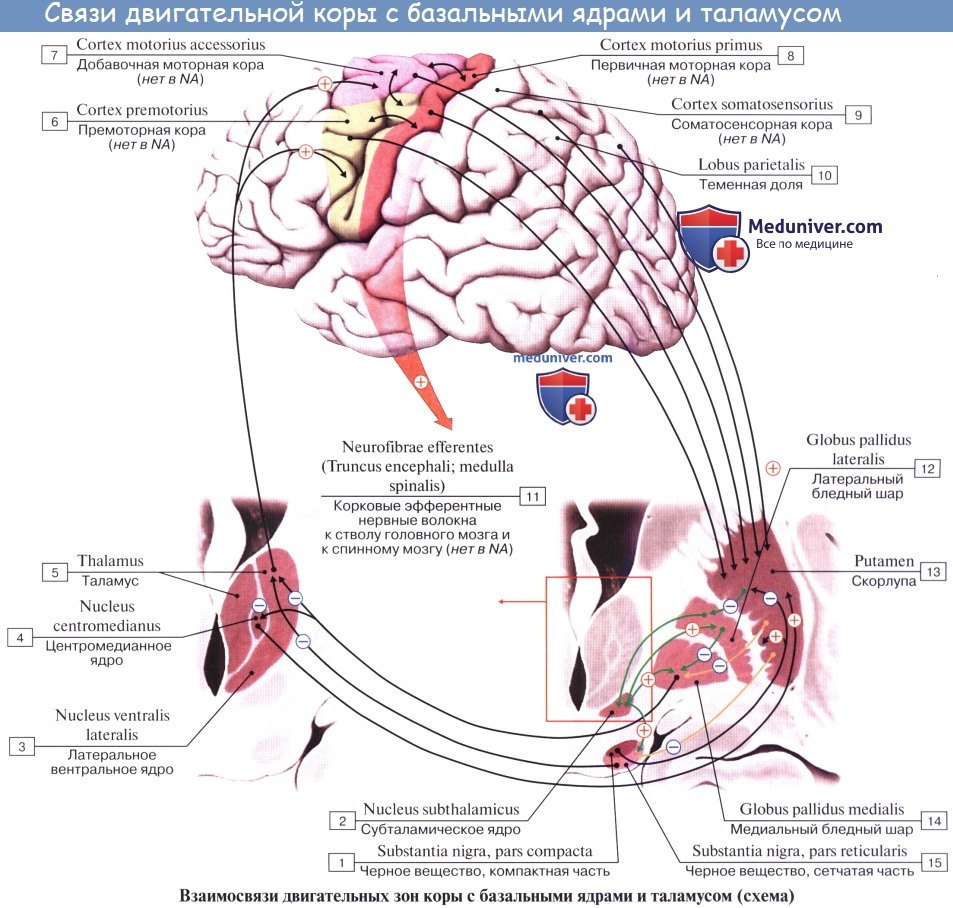
Таким образом, thalamus является подкорковым центром почти всех видов чувствительности. Отсюда чувствительные пути идут частью в подкорковые ядра (благодаря чему таламус является чувствительным центром экстрапирамидной системы), частью — непосредственно в кору (tractus thalamocorticalis).
Видео урок для зубрешки анатомия промежуточного мозга и таламической области
Редактор: Искандер Милевски. Дата последнего обновления публикации: 14.8.2020
Физиология человека и животных
Разделы
9. Промежуточный мозг. Роль ядер таламуса в передаче сигналов с периферии в кору больших полушарий. Гипоталамус как интегративно координирующий вегетативный центр мозга
Промежуточный мозг включает в себя таламус, эпиталамус (эпифиз) и гипоталамус и образует стенкиIIIжелудочка. Промежуточный мозг регулирует сложные двигательные рефлексы, координирует работу внутренних органов и осуществляет гуморальную регуляцию (обмен веществ, потребление воды и пищи, поддержание температуры тела). Вместе с большими полушариями промежуточный мозг участвует в организации всех сложных форм поведения, регуляции вегетативных реакций, то есть он интегрирует сенсорные, двигательные и вегетативные функции, обеспечивая деятельность организма как единого целого.
Таламус (зрительный бугор) представляет собой комплекс ядер в промежуточном мозге (у человека примерно 60 ядер), образующих множество прямых и обратных связей с корой больших полушарий. В таламусе осуществляется анализ афферентных сигналов практически от всех чувствительных рецепторов (кроме обонятельных), организация интегративных процессов, необходимых для регуляции функционального состояния и высшей нервной деятельности.
Ядра таламуса делят на две группы – специфические и неспецифические. Все ядра таламуса в разной степени обладают тремя общими функциями – переключающей, интегративной и модулирующей.
Специфические ядра таламуса:
1) Переключающие ядра:
1а) сенсорные – передают афферентную (чувствительную) информацию в сенсорные зоны коры;
1б) несенсорные – переключают в кору несенсорную импульсацию из разных отделов головного мозга (например, лимбические ядра таламуса).
2) Ассоциативные ядра – принимают импульсацию от других ядер таламуса. Благодаря их деятельности осуществляется объединение деятельности таламических ядер и различных зон ассоциативной коры.
Неспецифические ядра действуют как объединяющие посредники между стволом мозга и мозжечком, с одной стороны, и новой корой, лимбической системой и базальными ганглиями, с другой стороны, объединяя их в единую функциональную систему. Они обеспечивают модулирование, плавную настройку функционирования ЦНС. По своему функциональному значению они сходны с ретикулярной формацией. Но если ретикулярная формация осуществляет длительную и медленную активацию коры больших полушарий, то неспецифические ядра таламуса – быструю и кратковременную активацию.
Гипоталамус является вентральной частью промежуточного мозга. Макроскопически он включает в себя преоптическую область и область перекреста зрительных нервов, серый бугор и воронку, сосцевидные тела. Микроскопически в гипоталамусе выделяют около 50 пар ядер, которые топографически объединяют в 5 групп. Ядра гипоталамуса имеют мощное кровоснабжение.
Функции гипоталамуса:
Особое место в функциях гипоталамуса занимает регуляция деятельности гипофиза. Благодаря гипоталамо-гипофизарным связям гипоталамус является высшим центром эндокринной регуляции.
В промежуточном мозге выделяют таламус и гипоталамус
а) Гипоталамический контроль гипофиза. Кровоснабжение гипофиза осуществляют гипофизарные ветви внутренней сонной артерии. Одна часть ветвей снабжает капиллярную сеть в стенке воронки гипоталамуса. Отток от этих капилляров происходит в воротные (портальные) сосуды, проходящие в аденогипофизе (передней доле). Там они формируют вторичную капиллярную сеть, омывающую эндокринные клетки и впадающую в пещеристый синус.
Нейрогипофиз непосредственно кровоснабжают нижние гипофизарные артерии. Отток от капилляров происходит в пещеристый синус, через который секретированные гормоны из передней и задней долей попадают в общий кровоток.
Секрецию гипофиза контролируют два типа нейроэндокринных клеток. Нейроэндокринные клетки представляют собой истинные нейроны, так как имеют аксоны и дендриты, а также способны к проведению нервных импульсов. Их также считают истинными эндокринными клетками, поскольку они высвобождают свой секрет в капиллярную сеть. Секрет (за одним исключением, описанным ниже) представляет собой пептиды, синтезированные в скоплениях гранулярной эндоплазматической сети и упакованные в комплексах Гольджи. Эти пептиды прикреплены к длинноцепочечным полипептидам — нейрофизинам. Фенестрированная (пористая) капиллярная сеть расположена за пределами гематоэнцефалического барьера.
Тела нейроэндокринных клеток расположены в гипофизотропной зоне в нижней половине предоптической и бугорной областей. К соответствующим ядрам относят предоптическое, супраоптическое, паравентрикулярное, вентромедиальное и дугообразное (воронкообразное). Здесь можно выделить два типа нейронов: мелкоклеточные нейроны, расположенные до области срединного возвышения, и крупноклеточные нейроны, находящиеся до области задней доли гипофиза.
1. Мелкоклеточная нейроэндокринная система. Мелкоклеточные нейроны гипофизотропной зоны дают начало тубероинфундибулярному пути, который достигает капиллярной сети воронки гипоталамуса. Потенциалы действия, проходящие по этим нейронам, инициируют кальций-зависи-мый экзоцитоз выделяемых гормонов из одной группы нейронов и торможение высвобождения гормонов из другой, переносимых к аденогипофизу через портальные сосуды. Клетки аденогипофиза, продуцирующие рилизинг-гормоны (либерины, стимулируют выработку гормонов) и статины (угнетают выработку гормонов), представлены в таблице ниже. Единственный небелковый гормон мелкоклеточных клеток в левой колонке — пролактоста-тин (дофамин), секретируемый дугообразным (инфундибулярным) ядром.
Стимулирующие/ингибирующие гормоны не обладают строгой специфичностью: обычно основное их воздействие опосредовано на один тип клеток и дополнительно — на один или два других типа.
Мелкоклеточные нейроны гипофизотропной зоны имеют множественную регуляцию. К контролирующим факторам относят: деполяризацию восходящих волокон, восходящих из лимбической системы и ретикулярной формации; гиперполяризацию клетками локальных нейронных сетей с гамма-аминомасляной кислотой (ГАМК), часть из которых чувствительны к циркулирующим гормонам; торможение выделения нейромедиатора многочисленными опиатергическими вставочными нейронами промежуточной зоны гипоталамуса. Ситуация еще больше осложняется тем, что опиаты и другие модулирующие пептиды могут попадать в портальные сосуды и активировать рецепторы эндокринных клеток аденогипофиза. Стресс приводит к повышению секреции адренокортикотропного гормона (АКТГ), который, в свою очередь, стимулирует кору надпочечников с повышением в плазме концентрации глюкокортикоидов, в том числе и кортизола.
В норме кортизол по механизму отрицательной обратной связи возбуждает тормозные гипоталамические нейроны, имеющие глюкокортикоидные рецепторы. У пациентов, страдающих большим депрессивным расстройством, происходит нарушение этой обратной связи.
Нейроэндокринные клетки гипоталамуса. Кровоснабжение гипофиза, включая нейроэндокринные клетки аденогипофиза.
2. Крупноклеточная нейроэндокринная система. Крупноклеточные нейроны в супраоптическом и паравентрикулярном ядрах дают начало гипоталамо-гипофизарнму пути (супраоптикогипофизарному пути), нисходящему в нейрогипофиз (заднюю долю). Небольшая часть волокон входит в этот путь от опиатергических и других пептидергических нейронов перивентрикулярной области гипоталамуса, а также от аминергических нейронов ствола мозга.
В супраоптическом и паравентрикулярном ядрах отдельные группы нейронов секретируют два гормона: антидиуретический гормон (АДГ, вазопрессин) и окситоцин. Аксональные глыбки, содержащие секреторные гранулы с этими гормонами, составляют около половины объема нейрогипофиза. Самые крупные глыбки, называемые тельцами Херринга, могут достигать размеров эритроцита. Тельца Херринга обеспечивают локальный запас гранул, высвобождаемых более мелкими терминальными глыбками в капиллярное русло.
— Антидиуретический гормон. АДГ длительно стимулирует реабсорбцию воды в дистальных отделах извитых канальцев и собирательных трубочках почек. Основной регулятор электрической активности АДГ-секретирующих нейронов — осмотическое давление крови. Подъем осмотического давления даже на 1 % приводит к разбавлению крови до нормального уровня путем увеличения реабсорбции воды. Сами нейроны чувствительны к изменениям осмолярности, однако их чувствительность усиливают осморецепторы и волюморецепторы других отделов, в частности сосудистого и субфорникалъного околожелудоч-ковых органов.
Часть АДГ-секретирующих нейронов синтезирует также кортикотропин рилизинг-гормон (КРГ), оба гормона выделяются из коллатеральных ветвей в капиллярное русло воронки гипоталамуса. Интересно, что активность АДГ-секретирующих нейронов усиливается в состоянии стресса, и выброс АКТГ увеличивается при наличии АДГ в аденогипофизе. Торможение секреции АДГ приводит к несахарному диабету.
— Окситоцин. Основная функция окситоцина — участие в нейрогуморальном рефлексе, когда младенец присасывается к груди. Восходящая дуга рефлекса образована волокнами, идущими от соска к гипоталамусу по спиноретикулярному пути. Окситоцин высвобождают крупноклеточные нейроны в ответ на сосание. Попав в общий кровоток, он вызывает выделение молока путем возбуждения миоэпителиальных клеток, окружающих млечные протоки молочной железы.
Окситоцин также оказывает умеренное возбуждающее действие на мышцы матки в период родов. Афферентные импульсы в этом случае проходят по генитальному пути сразу после начала родов. Было обнаружено, что окситоцин и вазопрессин участвуют также в процессах обучения, сексуального и материнского поведения, а также при беспокойстве и агрессии.
б) Другие взаимосвязи и функции гипоталамуса. Гипоталамус непосредственно и косвенно осуществляет координацию нескольких различных функций, участвующих в поддержании гомеостаза. К этим функциям относят вегетативную регуляцию, терморегуляцию, регуляцию осмотического давления, сексуальное поведение, реакцию на стресс и цикл сонбодрствование. В некоторых ситуациях при изолированных поражениях возможно выпадение только одной функции, однако обычно развиваются более сложные функциональные нарушения. Таким образом, ядра, окружающие третий желудочек (перивентрикулярные), выполняют нейроэндокринную функцию; медиальные ядра отвечают за терморегуляцию, осморегуляцию и реакцию на стресс; латеральные ядра регулируют цикл сонбодрствование, пищевое поведение и жажду.
1. Вегетативные центры. Стимуляция передней гипоталамической области у животных приводит к развитию парасимпатических реакций: замедлению работы сердца, сужению зрачков, секреции слюны и усилению перистальтики кишечника. Напротив, стимуляция задней гипоталамической области приводит к симпатическим реакциям: усилению сердечного ритма и повышению артериального давления, расширению зрачков, торможению перистальтики кишечника. Волокна обоих отделов направляются к вегетативным ядрам ствола мозга и спинного мозга. В среднем мозге и мосту эти проводящие пути располагаются в заднем продольном пучке.
2. Терморегуляция. Предоптическое ядро переднего гипоталамуса содержит термочувствительные нейроны, которые запускают соответствующие реакции в ответ на изменения внутренней температуры тела. Возбуждение этих нейронов усиливают импульсы, полученные (через спиноретикулярный путь) от термочувствительных нейронов, иннервирующих кожу.
Внутренняя температура тела поддерживается с помощью механизмов, скоординированных передним гипоталамусом. Повышение базальной температуры может быть скорректировано ядром гипоталамуса, от которого отходят волокна, образующие синапсы с преганглионарными грудопоясничными нейронами бокового рога спинного мозга, усиливающими кровоток в коже и стимулирующими потовые железы. Одновременно происходит торможение механизмов теплопродукции в заднем гипоталамусе.
Гипоталамический контроль симпатической нервной системы с возрастом ослабевает. Этим объясняют тот факт, что пожилые люди особенно часто подвержены развитию гипотермии при холодной погоде.
Гипертермия—характеристика лихорадки. В ответ на инфекционные агенты (бактерии, вирусы, паразиты) тканевые макрофаги высвобождают эндогенные пирогены — белки, вызывающие смещение вверх значений «гипоталамического термостата». (Пирогены осуществляют этот процесс с помощью активации продукции простагландинов в гипоталамусе.) Основные механизмы повышения температуры тела до нового уровня — вазоконстрикция сосудов кожи и дрожь.
3. Жажда. Главным центром, контролирующим потребление жидкости, вероятно, служит медиальное предоптическое ядро, которое обрабатывает информацию от периферических рецепторов, регистрирующих объем циркулирующей крови и артериальное давление, снижение скорости кровотока, подъем уровня гормона ангиотензина (субфорникальный орган), а также изменения осмолярности (сосудистый орган терминальной пластинки) Эта информация поступает в кору больших полушарий, которая активирует модель поведения, направленную на удовлетворение потребности (например, чувство жажды).
4. Пищевое поведение. На пищевые привычки, очевидно, влияют социальные и культурные традиции, поэтому пищевые пристрастия широко варьируют как среди отдельных людей, так и обществе. Дугообразное ядро гипоталамуса обрабатывает поступающие сигналы, связанные с пищей, в форме взаимодействия между латеральным и вентромедиальным ядрами, которые вместе определяют базовый уровень требуемых калорий и питательных веществ, составляющий аттестат (заданный уровень аппетита). Дугообразное ядро чувствительно к уровню глюкозы и различным секретируемым пептидам, стимулирующим пищевое поведение (грелин вырабатывается желудком и стимулирует пищевое поведение; лептин секретируется адипоцитами и подавляет голод). Разрушение латерального гипоталамуса, или «центра питания», у кошки или крысы заставляет животное отказаться от еды.
Напротив, поражения вентромедиального отдела гипоталамуса, или «центра насыщения», у животных приводит к постоянному перееданию и выраженному ожирению. Интересен тот факт, что серотонин способен изменять значения аттестата, подавляя латеральное ядро. Люди, страдающие анорексией, обычно имеют повышенный уровень секреции серотонина, а страдающие булемией — пониженный.
5. Гипоталамическая реакция на психологический стресс. Стрессовое событие (психологическое, физическое или физиологическое) нарушает нормальный гомеостаз, и физиологические системы предпринимают попытку восстановления дисбаланса. Гипоталамус, а особенно гипоталамо-гипофизарно-надпочечниковая система (ГГНС) — неотъемлемая часть этого восстановительного механизма.
Паравентрикулярное ядро получает информацию от структур ствола мозга, реагирующих на различные физиологические стрессовые факторы, а также от лимбической системы, участвующей в эмоциональных реакциях. Кортикотропин-РГ, выделяемый паравентрикулярным ядром (действие усиливается одновременным высвобождением вазопрессина), приводит к секреции АКТГ аденогипофизом. АКТГ стимулирует выделение кортизола корой надпочечников. Кортизол, в свою очередь, активирует запасание энергии во всем организме.
Вероятно, имеются гендерные различия в типе физиологической и биологической реакции мужчин и женщин на стресс. При функциональной магнитно-резонансной томографии (фМРТ) у мужчин была обнаружена активация латеральной предлобной (префронтальной) коры (важного центра принятия решений в ответ на проблему или лишение), тогда как у женщин преобладала активация поясной извилины — основного коркового центра эмоционального контроля. Можно ли связать сложные программы поведения с изменениями, выявляемыми при фМРТ, остается неясным.
6. Злость и страх. Латеральное и вентромедиальное ядра отвечают за настроение, а также за пищевые привычки. Кошки, страдающие ожирением, проявляют высокую агрессивность вследствие поражения вентромедиального ядра. Напротив, животные, у которых путем стимуляции вентромедиального ядра развился недостаток веса, отличаются чрезмерной покорностью.
7. Сон и бодрствование. Гипоталамус играет ключевую роль как в возбуждающей системе мозга, так и в цикле сон-бодрствование. Мельчайшее (0,26 мм 3 ) надперекрестное ядро, расположенное на верхней поверхности зрительного перекреста, получает информацию непосредственно от сетчатки и служит водителем циркадного ритма мозга. Оно участвует в установлении нормального цикла сон-бодрствование посредством влияния на эндокринную, вегетативную и поведенческую функции (например, путем воздействия на эпифиз и секрецию им мелатонина).
Поражения задней области гипоталамуса могут сопровождаться гиперсомнией или даже комой. В этой области расположено серобугорно-сосцевидное ядро, содержащее сотни гистаминергических нейронов, аксоны которых широко распространяются по серому веществу головного и спинного мозга. Часть волокон проходит в ростральном направлении через медиальный пучок переднего мозга в сопровождении аминергических волокон от ствола мозга. Гистаминергические волокна, идущие к коре больших полушарий, широко разветвляются ниже колена мозолистого тела. Они разветвляются в поверхностных слоях лобной коры и направляются кзади — к теменной, затылочной и височной долям.
Получены физиологические данные в пользу выполнения гистаминергической системой функции возбуждения у животных. Серобугорно-сосцевидное ядро в норме активировано в состоянии бодрствования белком орексином, выделяемым небольшой группой нейронов латерального гипоталамуса. Нарушение продукции орексина может лежать в основе приступов засыпания при нарколепсии.
8. Сексуальное возбуждение. Подгруппа нейронов (третье интерстициальное ядро переднего гипоталамуса, ИЯПГ3) в медиальном отделе предоптического ядра более чем в два раза крупнее у мужчин, чем у женщин. Оно также богато андрогеновыми рецепторами и активируется циркулирующим в крови тестостероном. У женщин богатые эстрогеновыми рецепторами нейроны содержатся в вентромедиальном ядре. Электрическая стимуляция этих ядер у лабораторных животных приводила к развитию соответствующих половых реакций, что позволило доказать их аналогичную функцию у людей.
9. Память. Сосцевидные тельца относят к лимбическому кругу (Пейпса), включающему свод мозга, посылающий к нему волокна, и сосцевидно-таламический путь, оканчивающийся в переднем ядре таламуса. Этот круг участвует в процессах памяти.
в) Большое депрессивное расстройство. Большое депрессивное расстройство — состояние, сопровождающееся снижением настроения и развивающееся без видимых внешних причин. Заболевание встречают примерно у 4% взрослого населения, оно имеет генетическую предрасположенность: около 20% родственников первого порядка также страдают этим заболеванием. Фазы депрессии можно наблюдать уже в детском или подростковом возрасте.
Большое депрессивное расстройство характеризуется как минимум несколькими из нижеперечисленных признаков:
• Сниженный фон настроения с потерей интереса к повседневной деятельности и внешним событиям.
• Упадок сил, быстрая утомляемость, потеря аппетита и снижение либидо, запор.
• Снижение самооценки с чувством собственной неполноценности.
• Нарушение цикла сон-бодрствование, проявляющееся ранним пробуждением по утрам.
• Болезненность и боли. Рецидивирующие боли в животе могут имитировать органное заболевание.
• Периоды тревожности и беспокойства, а также склонность к суициду.
Применение моноаминов вначале было основано на случайных наблюдениях, при которых использование резерпина для лечения артериальной гипертензии приводило в качестве побочного эффекта к депрессии. Резерпин истощает депо моноаминов (серотонина, норадреналина и дофамина).
Вышеперечисленные симптомы характерны также для хронического стресса. Именно поэтому у пациентов с депрессией имеет место гиперактивация коры надпочечников, сывороточная концентрация кортизола повышена. Как уже отмечено, повышение сывороточной концентрации кортизола в норме тормозит продукцию кортиколиберина в гипоталамусе. У пациентов с депрессией отмечают относительную резистентность глюкокортикоидных рецепторов. Происходящие изменения лежат в основе теста подавления дексаметазоном. Дексаметазон—мощный синтетический глюкокортикоид, подавляющий секрецию АКТГ у здоровых людей.
Отростки части кортиколиберин-продуцирующих нейронов проходят непосредственно в мозг. В среднем мозге кортиколиберин тормозит мезокортикальные дофаминергические нейроны, которые в норме отвечают за положительную мотивацию. Кроме того, в среднем мозге он тормозит серотонинергические нейроны шва, играющие ключевую роль в циркадных ритмах в основном за счет обильной иннервации надперекрестного ядра.
К первой линии терапии относят препараты, улучшающие серотонинергиче-скую передачу. Широкий диапазон антидепрессантов имеет разные механизмы действия: часть из них, например, тормозит обратный захват из межсинаптической щели, другие тормозят разрушение моноаминоксидазой. Эффект наступает через несколько недель; латентный промежуток обусловлен десенситизацией (резистентностью) ауторецепторов на мембранах серотонинергических нейронов.
Электрошоковая терапия (ЭШТ)—наименее эффективный из антидепрессантов. Она, вероятно, приводит к десенситизации ауторецепторов и сенситизации (возбуждению) серотониновых рецепторов на клетках-мишенях и торможению норадренергической передачи.
г) Болезни гипоталамуса. Наиболее тяжелое расстройство функции гипоталамуса—несахарный диабет, развивающийся при повреждении гипоталамо-гипофизарных проводящих путей (иногда при опухоли в этой области или черепно-мозговой травме). Пациент пьет более 10 л воды в день и выделяет аналогичное количество мочи. Исторически термин insipidus указывал на отсутствие вкуса мочи («безвкусный») в противоположность сахарному диабету (diabetes mellitus), при котором моча имеет сладкий (mellitus) вкус вследствие содержания в ней сахара.
При гипофизэктомии (хирургическом удалении гипофиза), которую выполняют при лечении других заболеваний, вызывая лишь временный несахарный диабет, проводят рассечение воронки гипофиза в нижнем отделе. В течение короткого периода времени в капиллярную сеть срединного возвышения начинает секретироваться достаточное количество АДГ, обеспечивая адекватное сохранение жидкости.
В литературе описано широкое разнообразие поражений гипоталамуса. Причины также могут быть различны: опухоли, врожденные мальформации, черепно-мозговая травма. Клиническая картина сопровождается выраженным ожирением, вегетативными нарушениями, избыточной сонливостью и потерей памяти.
Околожелудочковые органы.
д) Околожелудочковые органы. Шесть зон мозга, прилежащих к системе желудочков, содержат нейроны и специализированные глиальные клетки, окружающие фенестрированные (пористые) капилляры. Их называют окопожелудочковыми органами (ОЖО) Срединное возвышение и нейрогипофиз описаны в основном тексте Сосудистый орган терминальной пластинки и субфорникальный орган, прилежащие к межжелудочковому отверстию, отдают волокна к супраоптическому и паравентрикулярному ядрам гипоталамуса и облегчают деполяризацию нейронов, секретирующих АДГ. При развитии гиповолемии почки секретируют ренин, который опосредует образование ангиотензина II, возбуждающего эти два ОЖО, завершая положительную обратную связь.
Эпифиз синтезирует мелатонин—гормональный амин, участвующий в цикле сон-бодрствование. Мелатонин синтезируется из серотонина, необходимые для этого ферменты имеются только в этой железе. Мелатонин высвобождается из капиллярного русла эпифиза ночью и оказывает снотворный эффект; к другим действиям относят очищение от патогенных свободных радикалов, выделяющихся из тканей в процессе их старения. Дневную секрецию подавляет активность симпатических волокон, подходящих к эпифизу от верхних шейных ганглиев по стенкам прямого венозного синуса. Важный центральный проводящий путь подходит из парных надперекрестных ядер через задний продольный пучок.
Начиная с третьего десятилетия жизни, в астроцитах эпифиза могут накапливаться известковые отложения («Эпифизарный песок»). Кальцификацию часто можно обнаружить на рентгенограммах черепа в прямой проекции. Смещение железы может указывать на новообразование в полости черепа. Однако и в норме эпифиз иногда немного отклонен влево, так как правое полушарие головного мозга на этом уровне обычно немного шире, чем левое.
Самое заднее поле (area postrema) погружено в крышу четвертого желудочка на уровне задвижки. Это хеморецепторная триггерная зона, или рвотный центр. Рвотный центр образован нейронами, чувствительными к широкому диапазону токсических веществ, и выполняет защитную функцию путем рефлекторного вызова рвоты через связи с гипоталамусом и ретикулярной формацией.
е) Резюме. Гипоталамус—двустороннее образование, расположенное за третьим желудочком. В сагиттальной плоскости его разделяют на переднюю (супраоптическую) область, включающую три ядра, среднюю (бугорную), содержащую пять ядер, и заднюю (сосцевидную) с тремя ядрами. Во фронтальной проекции гипоталамус разделяют на латеральную, медиальную и перивентрикулярную области.
Гипофиз контролируют нейроэндокринные клетки гипоталамуса, способные как к передаче импульсов, так и к секреции гормонов в капиллярное русло. Мелкоклеточные нейроэндокринные клетки расположены около срединного возвышения. Там они секретируют рилизинг-гормоны/статины в капиллярное русло, откуда они направляются к аденогипофизу по системе портальных сосудов. Крупные нейроэндокринные клетки образуют гипоталамо-гипофизарный путь, через который в капиллярное русло нейрогипофиза высвобождаются АДГ и окситоцин.
К околожелудочковым органам, расположенным за пределами гематоэнцефалического барьера, относят: срединное возвышение и нейрогипофиз; сосудистый орган терминальной пластинки и субфорникальный орган (оба органа участвуют в механизме обратной связи при регуляции объема плазмы); эпифиз, секретирующий мелатонин; самое заднее поле; субфорникальный орган.
В переднем и заднем отделах гипоталамуса расположены нейроны, возбуждающие парасимпатическую и симпатическую нервную систему соответственно. Органы терморегуляции поддерживают заданную температуру тела в основном за счет симпатической регуляции.
Стимуляция латеральной зоны гипоталамуса приводит к повышению потребления жидкости и пищи. Разрушение этой области или стимуляция вентромедиального центра насыщения приводит к отказу от пищи.
Редактор: Искандер Милевски. Дата публикации: 21.11.2018
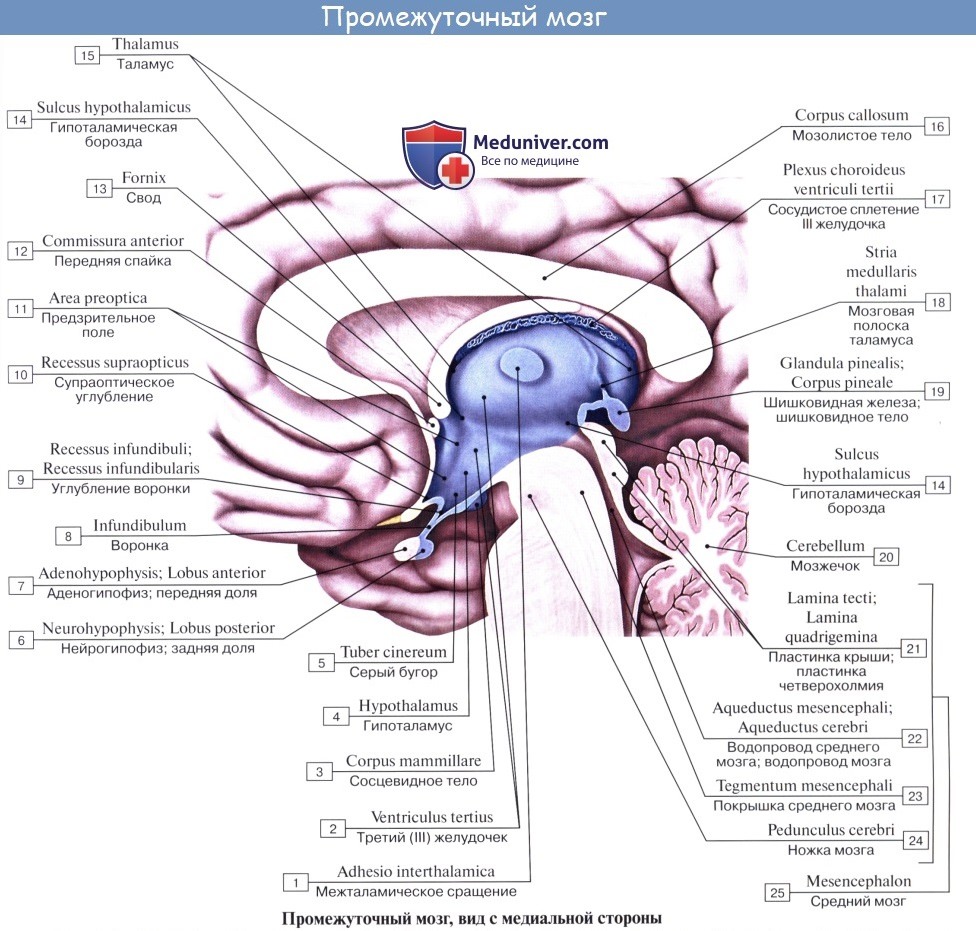
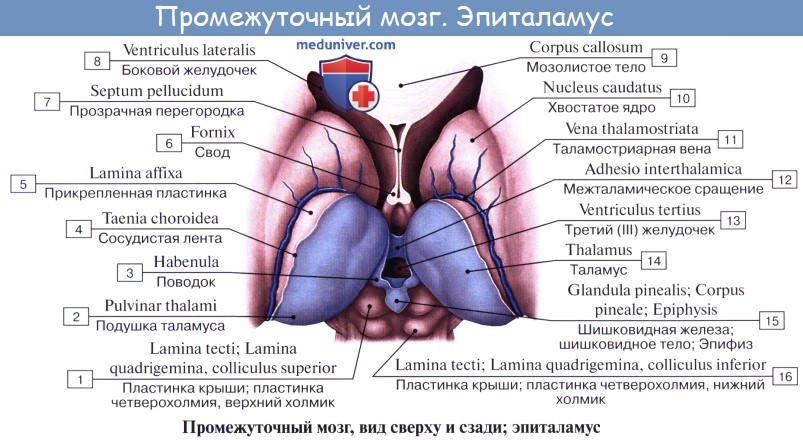
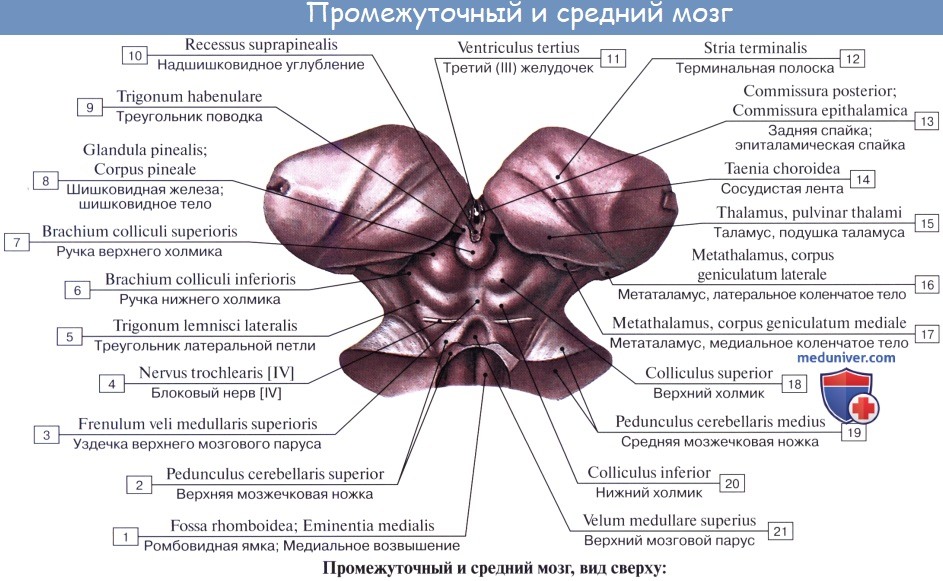
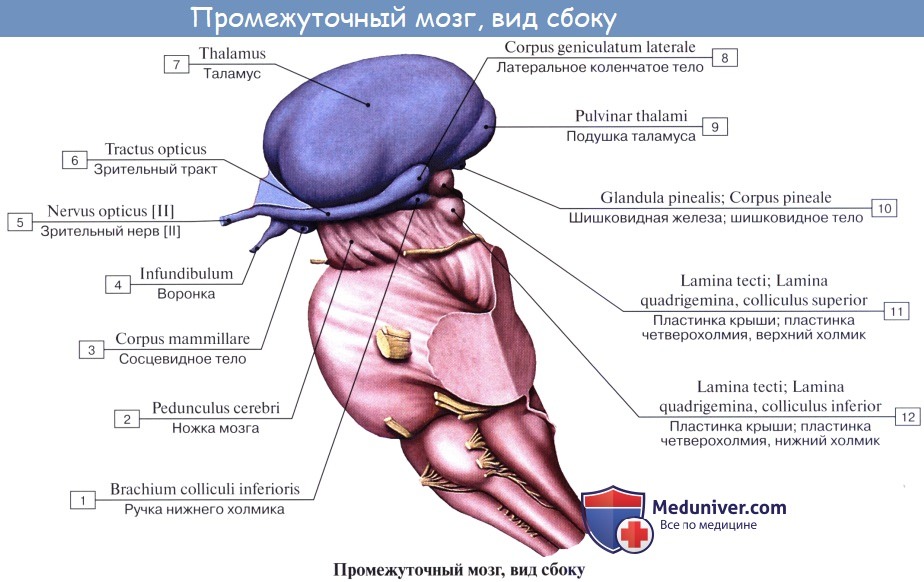
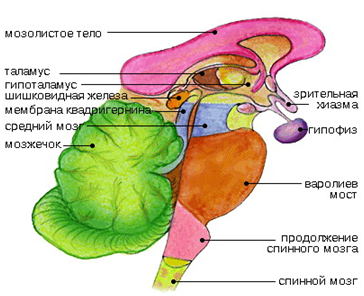 Промежуточный мозг включает в себя таламус, эпиталамус (эпифиз) и гипоталамус и образует стенки III желудочка. Промежуточный мозг регулирует сложные двигательные рефлексы, координирует работу внутренних органов и осуществляет гуморальную регуляцию (обмен веществ, потребление воды и пищи, поддержание температуры тела). Вместе с большими полушариями промежуточный мозг участвует в организации всех сложных форм поведения, регуляции вегетативных реакций, то есть он интегрирует сенсорные, двигательные и вегетативные функции, обеспечивая деятельность организма как единого целого.
Промежуточный мозг включает в себя таламус, эпиталамус (эпифиз) и гипоталамус и образует стенки III желудочка. Промежуточный мозг регулирует сложные двигательные рефлексы, координирует работу внутренних органов и осуществляет гуморальную регуляцию (обмен веществ, потребление воды и пищи, поддержание температуры тела). Вместе с большими полушариями промежуточный мозг участвует в организации всех сложных форм поведения, регуляции вегетативных реакций, то есть он интегрирует сенсорные, двигательные и вегетативные функции, обеспечивая деятельность организма как единого целого.
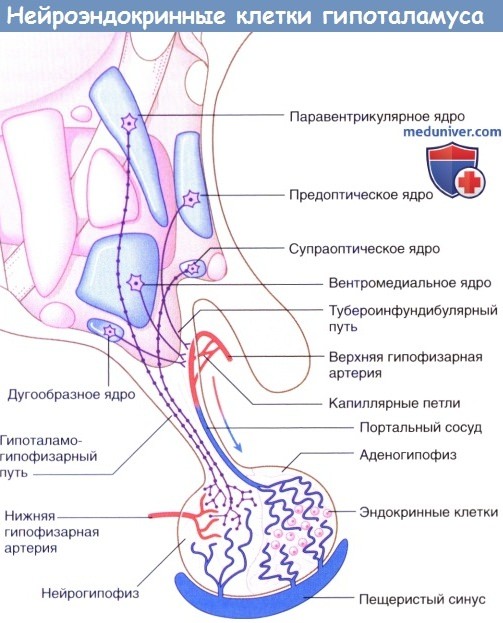 Нейроэндокринные клетки гипоталамуса.
Нейроэндокринные клетки гипоталамуса. 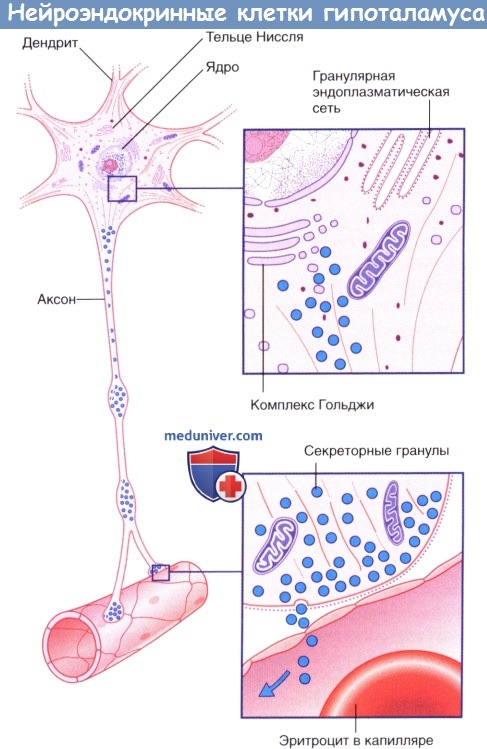 Морфология пептид-секретирующей нейроэндокринной клетки.
Морфология пептид-секретирующей нейроэндокринной клетки. 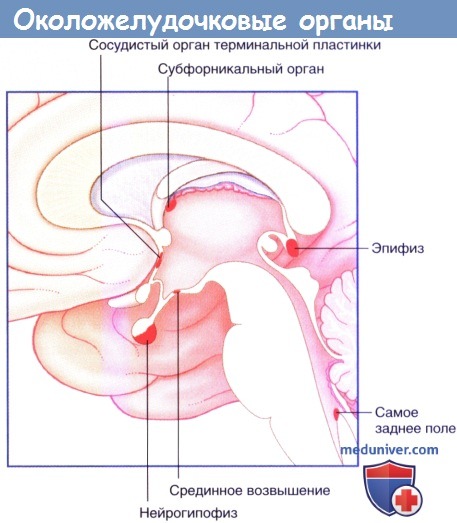 Околожелудочковые органы.
Околожелудочковые органы. 


