Триплоидия что это за болезнь
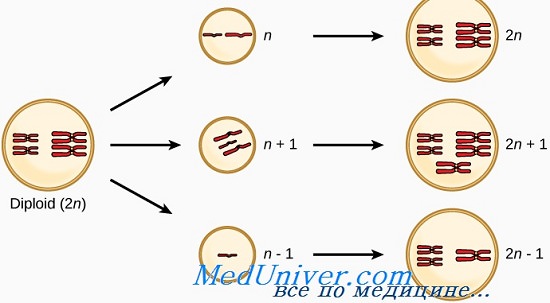
Набор хромосом с любым их количеством, кроме 46, называется гетероплоидным. Количество хромосом, кратное гаплоидному (b), — эуплоидное, любое другое число хромосом — анеуплоидное.
Триплоидия и тетраплоидия
Кроме диплоидного (2n) набора хромосом, характерного для нормальных соматических клеток, встречаются два других эуплоидных хромосомных набора, триплоидный (3n) и тетраплоидный (4n), изредка наблюдаемые в клиническом материале. Как триплоидия, так и тетраплоидия обнаружены у плодов; и хотя триплоидные младенцы могут родиться живыми, долго они не живут. Триплоидия встречается в 1-3% всех зачатий; большинство среди тех, которые донашиваются до конца I триместра, вызвано оплодотворением двумя сперматозоидами (диспермией).
Часть случаев также может быть вызвана нарушением одного из мейотических делений, приводящим к образованию диплоидной яйцеклетки или сперматозоида. Фенотипическая манифестация триплоидного кариотипа зависит от источника дополнительного набора хромосом; триплоиды с дополнительным набором отцовских хромосом обычно имеют аномальную плаценту и классифицируются как частично гидатиформные беременности (пузырный занос), с дополнительным набором материнских хромосом — спонтанно прерываются на более ранних сроках беременности. Тетраплоиды — всегда 92,ХХХХ или 92,XXYY; отсутствие половых конституций XXXY или XYYY подсказывает, что тетраплоидия происходит вследствие неправильного завершения первых делений зиготы.
Анеуплоидия
Анеуплоидии — наиболее частый клинически значимый тип хромосомных нарушений у человека, наблюдаемый, по крайней мере, в 5% клинически распознанных беременностей. Большинство анеуплоидных пациентов имеют или трисомии (три хромосомы вместо двух в норме), или, реже, моносомии (только одна конкретная хромосома). Как моносомии, так и трисомии бывают с серьезными фенотипическими последствиями.
Трисомия может захватывать любую часть генома, но трисомия целой хромосомы редко совместима с жизнью. Наиболее частый тип трисомии у живорожденных младенцев — трисомия 21 (кариотип 47.ХХ или XY.+21), хромосомная конституция, наблюдаемая у 95% пациентов с синдромом Дауна. Другие трисомии, встречающиеся у живорожденных детей, — трисомия 18 и трисомия 13. Примечательно, что все эти три (13, 18 и 21) аутосомы — с самым низким числом генов; возможно, трисомии аутосом с большим числом генов в большинстве случаев детальны.
Моносомия по целой хромосоме почти всегда летальна; важное исключение — моносомия хромосомы X, наблюдаемая при синдроме Тернера.
Хотя причины анеуплоидий до конца не понятны, известно, что наиболее частый механизм их образования — нерасхождение хромосом в мейозе. Это связано с неспособностью пары хромосом правильно разойтись в дочерние клетки в одном из делений мейоза, обычно в первом. Последствия нерасхождения в ходе первого и второго делений различны. Если ошибка происходит в течение первого деления мейоза, гамета с 24 хромосомами содержит в паре как отцовскую, так и материнскую хромосому.
Если же она происходит в ходе второго деления, гамета с дополнительной хромосомой содержит две одинаковые копии или отцовской, или материнской хромосомы. (Строго говоря, упомянутые утверждения относятся только к отцовской или материнской центромере, так как обычно в предыдущем делении мейоза между гомологичными хромосомами происходит рекомбинация, приводя к генетическим различиям между хроматидами и, таким образом, между соответствующими дочерними хромосомами).
Склонность хромосомной пары к нерасхождению сильно связана с отклонениями в частоте или положении рекомбинаций в первом делении мейоза. Хромосомная пара с малым числом рекомбинаций (или с их отсутствием), а также с рекомбинациями, расположенными слишком близко к центромере или теломере, более подвержена нерасхождению, чем хромосомная пара с типичным числом и распределением событий рекомбинации.
Дополнительно к классическому нерасхождению, когда неправильное разделение хромосом — результат нарушенного спаривания или рекомбинации, существует другой механизм, лежащий в основе анеуплоидий, — преждевременное разделение сестринских хроматид в мейозе I вместо мейоза II. Если это случается, разделенные хроматиды могут случайным образом расходиться в овоцит и полярное тельце, что также ведет к образованию несбалансированной гаметы.
Описаны также более сложные формы множественной анеуплоидий. Гаметы редко несут более одной дополнительной хромосомы. Однако если нерасхождение происходит сразу в двух делениях мейоза или одновременно как в мужских, так и в женских гаметах, это приводит к появлению чрезвычайно редко встречающихся зигот (за исключением половых хромосом) с необычным количеством хромосом. Нерасхождение также может происходить в митотическом делении уже после образования зиготы. Если подобное происходит в первых делениях зиготы, появляется клинически значимый мозаицизм.
В некоторых линиях злокачественных клеток и клеточных культурах митотическое нерасхождение может приводить к значительным аномалиям в кариотипе.
Большой вклад в диагностику анеуплоидий, особенно пренатальных, внесло использование многоцветной FISH в интерфазных клетках. Метод обеспечивает быструю диагностику без необходимости культивирования клеток. В настоящее время многие лаборатории пренатальной цитогенетики выполняют интерфазный анализ для выявления анеуплоидий хромосом 13, 18, 21, X и Y, т.е. тех пяти хромосом, которые участвуют в большинстве анеуплоидий у новорожденных.
Редактор: Искандер Милевски. Дата обновления публикации: 18.3.2021
Триплоидия
Синдром триплоидии – крайне редкое хромосомное расстройство. Лица с этим синдромом имеют в своих клетках по три (вместо двух) каждой хромосомы и в общей сложности их получается шестьдесят девять, а не нормальных сорок шесть. Почти всегда, для беременных плодом с этим синдромом, беременность заканчивается ранним выкидышем. Тем не менее, некоторые дети рождались и доживали до пяти месяцев.
Признаки Триплоидии
Эти дети, как правило, малы и имеют многочисленные врожденные дефекты. Те, которые выживают, как правило, имеют мозаичную форму этого синдрома, это означает, что одни клетки имеют нормальный набор (46) хромосом, а другие имеют дополнительный набор хромосом.Триплоидия развивается немного чаще у мальчиков чем у девочек
Пренатальная диагностика
1) Инвазивные исследования (амниоцентез, биопсия хориона) в основном назначают тем женщинам, у которых наблюдается повышенный риск того, что родится малыша с синдромом Эдвардса, например, пациенткам, чей возраст превышает 35 лет или с плохими результатами неинвазивных тестов: УЗИ и анализов. Инвазивные методы диагностики являются высокоточными, однако, учитывая риск осложнений, не подходят для массового проведения всем беременным, а проводятся только по особым показаниям.
2) Неинвазивные технологии, так называемые скрининги. Скрининг – комплексное исследование беременных женщин на наличие у плода хромосомных аномалий. Выделено несколько признаков, указывающих на высокий риск наличия заболевания, которые может выявить УЗИ плода (отсутствие носовой кости, увеличенная толщина воротникового пространства, недостаточная длина бедренных и плечевых костей и другие особенности). В комплексе с УЗИ идёт биохимический анализ крови матери на такие гормоны как свободный бета-ХГЧ и PAPP-A. Полученные данные по биохимическим маркерам анализируют в совокупности с результатами ультразвукового исследования, а результат всего скрининга представляет собой расчет риска наличия хромосомной аномалии у плода.
Однако при использовании стандартных тестов на синдром Эдвардса, лишь у 3% женщин, направленных на инвазивную диагностику действительно подтверждается наличие заболевания. В то же время не исключены и ложно-отрицательные результаты, когда скрининг показывает низкий риск, а ребенок рождается с хромосомной патологией.
Триплоидия
Синдром триплоидии – крайне редкое хромосомное расстройство. Лица с этим синдромом имеют в своих клетках по три (вместо двух) каждой хромосомы и в общей сложности их получается шестьдесят девять, а не нормальных сорок шесть. Почти всегда, для беременных плодом с этим синдромом, беременность заканчивается ранним выкидышем. Тем не менее, некоторые дети рождались и доживали до пяти месяцев. 
Признаки Триплоидии
Эти дети, как правило, малы и имеют многочисленные врожденные дефекты. Те, которые выживают, как правило, имеют мозаичную форму этого синдрома, это означает, что одни клетки имеют нормальный набор (46) хромосом, а другие имеют дополнительный набор хромосом.Триплоидия развивается немного чаще у мальчиков чем у девочек. 
Пренатальная диагностика
1) Инвазивные исследования (амниоцентез, биопсия хориона) в основном назначают тем женщинам, у которых наблюдается повышенный риск того, что родится малыша с синдромом Триплоидии, например, пациенткам, чей возраст превышает 35 лет или с плохими результатами неинвазивных тестов: УЗИ и анализов. Инвазивные методы диагностики являются высокоточными, однако, учитывая риск осложнений, не подходят для массового проведения всем беременным, а проводятся только по особым показаниям.
2) Неинвазивные технологии, так называемые скрининги. Скрининг – комплексное исследование беременных женщин на наличие у плода хромосомных аномалий. Выделено несколько признаков, указывающих на высокий риск наличия заболевания, которые может выявить УЗИ плода (отсутствие носовой кости, увеличенная толщина воротникового пространства, недостаточная длина бедренных и плечевых костей и другие особенности). В комплексе с УЗИ идёт биохимический анализ крови матери на такие гормоны как свободный бета-ХГЧ и PAPP-A. Полученные данные по биохимическим маркерам анализируют в совокупности с результатами ультразвукового исследования, а результат всего скрининга представляет собой расчет риска наличия хромосомной аномалии у плода.
Однако при использовании стандартных тестов на синдром Триплоидии, лишь у 3% женщин, направленных на инвазивную диагностику действительно подтверждается наличие заболевания. В то же время не исключены и ложно-отрицательные результаты, когда скрининг показывает низкий риск, а ребенок рождается с хромосомной патологией.
Причины генетического бесплодия

Согласно оценкам экспертов Всемирной организации здравоохранения, около 50 миллионов супружеских пар во всем мире (что составляет примерно 7% от их общего количества) не могут завести ребенка из-за мужского и/или женского бесплодия. На данный момент считается, что в 50% случаев причиной являются генетические нарушения.
Современная наука достигла больших успехов в изучении причин генетического бесплодия, особенно с появлением метода секвенирования нового поколения (next generation sequencing, NGS), позволяющего быстро и с минимальными трудозатратами «читать» последовательность ДНК. Тем не менее, этот вопрос остается сложным, и в имеющихся на данный момент знаниях остается еще немало пробелов.
Существует множество генов, влияющих на мужскую и женскую репродуктивную функцию. Например, в одних только яичках мужчины экспрессируется около 2300 генов. Наука продолжает развиваться, и список доступных анализов ежегодно пополняется новыми генетическими тестами.
Генетические изменения, способные приводить к невозможности зачатия и вынашивания беременности, бывают разными:
Хромосомные аномалии
Хромосомные нарушения представляют большой интерес в аспекте вопросов бесплодия, так как они ответственны более чем за половину всех выкидышей в первом триместре беременности. В большинстве случаев хромосомные аномалии не наследуются – они возникают случайно в половых клетках или клетках эмбриона. При этом происходят выкидыш, мертворождение, либо ребенок рождается с серьезными пороками развития.
В каждой клетке человеческого тела содержится 46 хромосом. В яйцеклетке и сперматозоиде их по 23 – соответственно, ребенок получает половину набора от матери и половину от отца. Состояние, при котором меняется количество хромосом, называется анеуплоидией. Некоторые примеры, связанные с бесплодием:
Помимо изменений количества хромосом, встречаются нарушения их структуры – аберрации.
Их основные разновидности:
Моногенные заболевания
Моногенные заболевания характеризуются возникновением мутации в одном гене, отвечающем за синтез определенного белка. Эти патологии передаются от родителей детям, причем типы наследования бывают разными.
В настоящее время известно много моногенных мутаций, связанных с бесплодием. Их список постоянно пополняется.
Мультифакториальные заболевания
Мультифакториальные, или полигенные заболевания имеют сложный патогенез. В их развитии принимают участие генетические нарушения (как наследственные, так и приобретенные), образ жизни, воздействия внешней среды. На «неправильные» гены накладываются такие факторы, как особенности питания, уровень физической активности, экологическая обстановка, вредные привычки, стрессы, прием различных лекарственных препаратов и пр.
Типичные примеры мультифакториальных заболеваний – сахарный диабет и сердечно-сосудистые патологии. Среди причин женского бесплодия важное значение имеют следующие заболевания:
Эпигенетические изменения
Эпигенетические изменения возникают в результате различных процессов, когда последовательность ДНК остается нормальной, но меняется активность генов. Например, это происходит в результате метилирования – прикрепления особых метильных групп к определенным участкам ДНК. Некоторые научные исследования показали, что эпигенетические механизмы играют роль в развитии бесплодия.

В каких случаях рекомендуется пройти генетическое тестирование?
Обычно врачи направляют пациентов на консультации к клиническим генетикам в следующих случаях:
Генетик собирает семейный анамнез, анализирует родословную и при необходимости назначает анализы. Для диагностики причин генетического бесплодия применяют разные методы: цитогенетический анализ, полимеразную цепную реакцию (ПЦР), флуоресцентную гибридизацию in situ (FISH), микрочипирование, секвенирование нового поколения.
Чтобы выявить распространенные аномалии у плода, на 16–20-й неделях беременности проводят тройной тест. По показаниям выполняют различные инвазивные исследования. Более современный и точный метод диагностики – неинвазивное пренатальное тестирование (НИПТ), во время которого изучают ДНК плода в крови матери.
Если у одного или обоих партнеров из пары, страдающей бесплодием, выявляют генетические нарушения, в ряде случаев могут помочь различные методы лечения или вспомогательные репродуктивные технологии (ЭКО, ИКСИ). Но иногда проблему не удается решить. В таких ситуациях врач предложит рассмотреть возможность использования донорских яйцеклеток или сперматозоидов.
С возрастом репродуктивные возможности уменьшаются, а в клетках накапливаются мутации, которые могут помешать наступлению и вынашиванию беременности, вызвать тяжелые патологии у ребенка. Поэтому женщинам, которые планируют забеременеть после 35 лет, стоит подумать о возможности сохранить свои яйцеклетки в криобанке. В дальнейшем ими можно воспользоваться в любое время – это будет своего рода «страховка» репродуктивной функции.
Если вы решили сохранить собственные половые клетки или воспользоваться донорскими, важно подобрать надежный банк половых клеток. На данный момент Репробанк является одним из крупнейших на территории России и СНГ. Наше криохранилище оснащено новейшим оборудованием, а в каталоге представлено большое количество доноров, среди которых любая пара наверняка сможет подобрать подходящего.

Зиновьева Юлия Михайловна
Ведёт генетическое обследование доноров Репробанка, осуществляет подбор доноров для пар, имеющих ранее рождённых детей с установленной генетической патологией.
Остановка развития зародыша
В основу статьи положены работы проф. Буэ.
Остановка развития зародыша в дальнейшем приводит к изгнанию плодного яйца, что проявляется в виде самопроизвольного выкидыша. Однако во многих случаях остановка развития происходит на очень ранних сроках и сам факт зачатия остается неизвестным для женщины. В большом проценте случаев такие выкидыши связаны с хромосомными аномалиями у зародыша.
Самопроизвольные выкидыши
Самопроизвольные выкидыши, определением которых служит «самопроизвольное прерывание беременности между сроком зачатия и сроком жизнеспособности плода», во многих случаях с большим трудом поддаются диагностике: большое число выкидышей происходит на очень ранних сроках: задержки месячных не происходит, или эта задержка настолько мала, что сама женщина не подозревает о беременности.
Клинические данные
Изгнание плодного яйца может произойти внезапно, или ему могут предшествовать клинические симптомы. Чаще всего угроза выкидыша проявляется кровянистыми выделениями и болями внизу живота, переходящими в схватки. Далее следуют изгнание плодного яйца и исчезновение признаков беременности.
Клиническое обследование может выявить несоответсвие между предполагаемым сроком беременности и размерами матки. Уровни гормонов в крови и моче могут быть резко снижены, указывая на отсутствие жизнеспособности зародыша. Ультразвуковое исследование позволяет уточнить диагноз, выявляя либо отсутствие зародыша («пустое плодное яйцо»), либо отставание в развитии и отсутствие сердцебиения
Клинические проявления самопроизвольного выкидыша значительно варьируют. В одних случаях выкидыш проходит незамеченным, в других — сопровождается кровотечением и может потребовать выскабливания полости матки. Хронология симптоматики может косвенно указывать на причину самопроизвольного выкидыша: кровянистые выделения с ранних сроков беременности, остановка роста матки, исчезновение признаков беременности, «немой» период в течение 4-5 недель, а затем изгнание плодного яйца чаще всего свидетельствуют о хромосомных нарушениях зародыша, а соответствие срока развития зародыша сроку выкидыша говорит в пользу материнских причин невынашивания беременности.
Анатомические данные
Анализ материала самопроизвольных выкидышей, сбор которого был начат в начале двадцатого века в Институте Карнеги, позволил выявить огромный процент аномалий развития среди абортусов ранних сроков
В 1943 году Хертиг и Шелдон опубликовали результаты патологоанатомического исследования материала 1000 выкидышей на ранних сроках. Материнские причины невынашивания беременности были ими исключены в 617 случаев. Современные данные указывают на то, что мацерированные зародыши во внешне нормальных оболочках тоже могут быть связаны с хромосомными аномалиями, что в сумме составляет около 3/4 всех случаев данного исследования.
| Морфологическое исследование 1000 абортусов (по Hertig и Sheldon, 1943) | ||
| Грубые патологические нарушения плодного яйца: плодное яйцо без зародыша или с недифференцированным зародышем | 489 | |
| Локальные аномалии зародышей | 32 | |
| Аномалии плаценты | 96 | 617 |
| Плодное яйцо без грубых аномалий | ||
| с мацерированными зародышами | 146 | |
| 763 | ||
| с немацерированными зародышами | 74 | |
| Аномалии матки | 64 | |
| Другие нарушения | 99 | |
Дальнейшие исследования Микамо и Миллера и Полланда позволили уточнить связь между сроком выкидыша и частотой нарушений развития зародыша. Оказалось, что чем меньше срок выкидыша, тем частота аномалий выше. В материалах выкидышей, происшедших до 5-й недели после зачатия макроскопические морфологические аномалии плодного яйца встречаются в 90% случаев, при сроке выкидыша от 5 до 7 недель после зачатия — в 60%, при сроке больше 7 недель после зачатия — менее, чем в 15—20%.
Важность значения остановки развития зародыша в ранних самопроизвольных выкидышах была показана прежде всего фундаментальными исследованиями Артура Хертига, который в 1959 г. опубликовал результаты исследования человеческих зародышей до 17 дней после зачатия. Это был плод его 25-летней работы.
У 210 женщин в возрасте до 40 лет, идущих на операцию гистерэктомии (удаления матки) дата операции была сопоставлена с датой овуляции (возможного зачатия). После операции матки подвергались самому тщательному гистологическому исследованию на предмет выявления возможной беременности малого срока. Из 210 женщин только 107 были оставлены в исследовании в связи с обнаружением признаков овуляции, и отсутствием грубых нарушений труб и яичников, препятствующих наступлению беременности. Было обнаружено 34 плодных яйца, из них 21 плодное яйцо было внешне нормальным, а 13 (38%) имело явные признаки аномалий, которые, по мнению Хертига, обязательно привели бы к выкидышу или на этапе имплантации или вскоре после имплантации. Поскольку в то время не было возможности проведения генетического исследования плодных яиц, причины нарушений развития зародышей оставались неизвестными.
При обследовании женщин с подтвержденной фертильностью (все пациентки имели по несколько детей) было обнаружено, что одно из трех плодных яиц имеет аномалии и подвергается выкидышу до появления признаков беременности.
Эпидемиологические и демографические данные
Нечеткая клиническая симптоматика ранних самопроизвольных выкидышей приводит к тому, что достаточно большой процент выкидышей на малых сроках проходит незамеченным женщинами.
В случае клинически подтвержденных беременностей около 15% всех беременностей заканчивается выкидышем. Большая часть самопроизвольных выкидышей (около 80%) происходит в первом триместре беременности. Однако если принять во внимание тот факт, что выкидыши часто случаются спустя 4-6 недель после остановки развития беременности, можно сказать, что с первым триместром связано более 90% всех самопроизвольных выкидышей.
Специальные демографические исследования позволили уточнить частоту внутриутробной смертности. Так, Френч и Бирман в 1953 — 1956 гг. регистрировали все беременности у женщин острова Канаи и показали, что из 1000 беременностей, диагностированных при сроке после 5 недель, 237 не увенчались рождением жизнеспособного ребенка.
Анализ результатов нескольких исследований позволил Леридону составить таблицу внутриутробной смертности, включающей в себя и неудачи оплодотворения (половой акт в оптимальные сроки — в течение суток после овуляции).
| Полная таблица внутри утробной смертности (на 1000 яйцеклеток, подвергшихся риску оплодотворения) (по Leridon, 1973) | ||
|---|---|---|
| Недели после зачатия | Остановка развития с последующим изгнанием | Процент продолжающихся беременностей |
| 16* | 100 | |
| 0 | 15 | 84 |
| 1 | 27 | 69 |
| 2 | 5,0 | 42 |
| 6 | 2,9 | 37 |
| 10 | 1,7 | 34,1 |
| 14 | 0,5 | 32,4 |
| 18 | 0,3 | 31,9 |
| 22 | 0,1 | 31,6 |
| 26 | 0,1 | 31,5 |
| 30 | 0,1 | 31,4 |
| 34 | 0,1 | 31,3 |
| 38 | 0,2 | 31,2 |
| * — неудачи зачатия | ||
Все эти данные указывают на огромную частоту самопроизвольных выкидышей и на важную роль нарушений развития плодного яйца в этой патологии.
Эти данные отражают общую частоту нарушений развития, не выделяя среди них конкретные экзо- и эндогенные факторы (иммунологические, инфекционные, физические, химические и т. д.).
Важно отметить, что независимо от причины повреждающего воздействия, при исследовании материала выкидышей обнаруживается очень большая частота генетический нарушений (хромосомных аберраций (на сегодня изучены лучше всего) и генных мутаций) и аномалий развития, как, например, дефекты развития нервной трубки.
Хромосомные аномалии, ответственные за остановку развития беременности
Цитогенетические исследования материала выкидышей позволили уточнить характер и частоту тех или иных хромосомных аномалий.
Общая частота
При оценке результатов больших серий анализов следует иметь в виду следующее. На результаты исследований подобного рода могут оказать значительное влияние следующие факторы: способ сбора материала, относительная частота более ранних и более поздних выкидышей, доля материала искусственных абортов в исследовании, часто не поддающаяся точной оценке, успех культивирования клеточных культур абортуса и хромосомного анализа материала, тонкие методы обработки мацерированного материала.
Общая оценка частоты хромосомных аберраций при невынашивании беременности составляет около 60%, а в первом триместре беременности — от 80 до 90%. Как будет показано ниже, анализ, основанный на стадийности развития зародыша, позволяет сделать гораздо более точные выводы.
Относительная частота
Практически все большие исследования хромосомных аберраций в материале выкидышей дали поразительно сходные результаты относительно характера нарушений. Количественные аномалии составляют 95% всех аберраций и распределяются следующим образом:
| трисомии | 50—55% |
| моносомии X | 15—20% |
| триплоидии | 15—20% |
| тетраплоидии | 5% |
Количественные хромосомные аномалии
Различные типы количественных хромосомных аберраций могут возникать в результате:
Моносомии
Моносомия X (45,X) представляет одну из часто встречающихся аномалий в материале самопроизвольных выкидышей. При рождении она соответствует синдрому Шерешевского-Тернера, и при рождении она встречается реже, чем другие количественные аномалии половых хромосом. Эта бросающаяся в глаза разница между относительно высокой частотой обнаружения лишних X-хромосом у новорожденных и относительно редким обнаружением моносомии X у новорожденных указывает на высокую частоту летальности моносомии X у зародыша. Кроме того, обращает на себя внимание очень большая частота мозаик у больных с синдромом Шерешевского-Тернера. В материале выкидышей, наоборот, мозаики с моносомией X крайне редки. Данные исследований показали, что только менее 1% всех моносомий X доходит до срока родов. Моносомии аутосом в материале выкидышей встречаются довольно редко. Это очень контрастирует с высокой частотой соответствующих трисомий.
Трисомии
В материале выкидышей трисомии представляют более половины всех количественных хромосомных аберраций. Обращает на себя внимание то, что в случаях моносомий недостающей хромосомой обычно оказывается X-хромосома, а в случаях избыточных хромосом, дополнительная хромосома чаще всего оказывается аутосомой.
Точная идентификация дополнительной хромосомы стала возможна благодаря методу G-бэндинга. Исследования показали, что все аутосомы могут принимать участие в нон-дисджанкшн (см. таблицу). Обращает на себя внимание, что три хромосомы, чаще всего встречающиеся при трисомиях новорожденных (15-я, 18-я и 21-я) чаще всего обнаруживаются и при летальных трисомиях у зародышей. Вариации относительных частот различных трисомий у зародышей отражают во многом сроки, на которых происходит гибель зародышей, поскольку, чем более летальной является комбинация хромосом, тем на более ранних сроках происходит остановка развития, тем реже будет обнаруживаться такая аберрация в материалах выкидышей (чем меньше срок остановки развития, тем труднее обнаружить такой зародыш).
| Лишняя хромосома при летальных трисомиях у зародыша (данные 7 исследований: Буэ (Франция), Карр (Канада), Кризи (Великобритания), Дилл (Канада), Кадзи (Швейцария), Такахара (Япония), Теркелсен (Дания)) | |||
|---|---|---|---|
| Дополнительная аутосома | Количество наблюдений | ||
| A | 1 | ||
| 2 | 15 | ||
| 3 | 5 | ||
| B | 4 | 7 | |
| 5 | |||
| C | 6 | 1 | |
| 7 | 19 | ||
| 8 | 17 | ||
| 9 | 15 | ||
| 10 | 11 | ||
| 11 | 1 | ||
| 12 | 3 | ||
| D | 13 | 15 | |
| 14 | 36 | ||
| 15 | 35 | ||
| E | 16 | 128 | |
| 17 | 1 | ||
| 18 | 24 | ||
| F | 19 | 1 | |
| 20 | 5 | ||
| G | 21 | 38 | |
| 22 | 47 | ||
Триплоидии
Крайне редко наблюдаемые при мертворождениях, триплоидии составляют пятую по частоте хромосомную аномалию в материале выкидыше. В зависимости от соотношения половых хромосом может быть 3 варианта триплоидий: 69XYY (самая редкая), 69, XXX и 69, XXY (самая частая). Анализ полового хроматина показывает, что при конфигурации 69, XXX чаще всего обнаруживается только одна глыбка хроматина, а при конфигурации 69, XXY чаще всего половой хроматин не обнаруживается.
Приведенный ниже рисунок иллюстрирует различные механизмы, приводящие к развитию триплоидии (диандрию, дигинию, диспермию). С помощью специальных методов (хромосомные маркеры, антигены тканевой совместимости) удалось установить относительную роль каждого из этих механизмов в развитии триплоидии у зародыша. Оказалось, что на 50 случаев наблюдений триплоидия была следствием дигинии в 11 случаях (22%), диандрии либо диспермии — в 20 случаях (40%), диспермии — в 18 случаях (36%).
| Механизмы образования триплоидной зиготы |
|---|
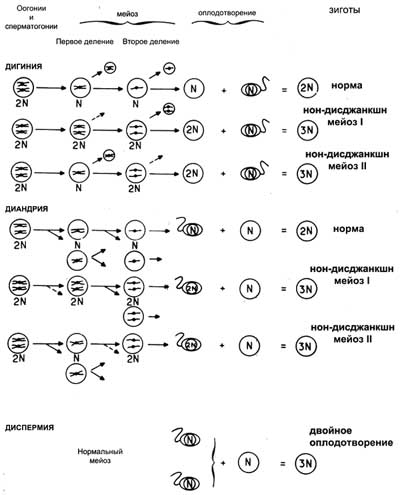 |
Тетраплоидии
Тетраплоидии встречаются примерно в 5% случаев количественных хромосомных аберраций. Чаще всего встречаются тетраплоидии 92, XXXX. Такие клетки всегда содержат 2 глыбки полового хроматина. В клетках с тетраплоидией 92, XXYY никогда не бывает видно полового хроматина, но в них обнаруживают 2 флуоресцирующие Y-хромосомы.
Двойные аберрации
Большая частота хромосомных аномалий в материале выкидышей объясняет высокую частоту комбинированных аномалий в одном и том же зародыше. Напротив, у новорожденных комбинированные аномалии крайне редки. Обычно в таких случаях наблюдаются комбинации аномалии половой хромосомы и аномалии аутосомы.
В связи с более высокой частотой аутосомных трисомий в материале выкидышей, при комбинированных хромосомных аномалиях у абортусов чаще всего встречаются двойные аутосомные трисомии. Трудно сказать, связаны ли такие трисомии с двойным «нон-дисджанкшн» в одной и той же гамете, или со встречей двух аномальных гамет.
Частота сочетаний различных трисомий в одной и той же зиготе носит случайный характер, что позволяет предположить независимость друг от друга появления двойных трисомий.
Комбинация двух механизмов, приводящих к появлению двойных аномалий, позволяет объяснить появление других аномалий кариотипа, встречающихся при выкидышах. «Нон-дисджанкшн» при образовании одной из гамет в сочетании с механизмами образования полиплоидии объясняет появление зигот с 68 или 70 хромосомами. Сбой первого митотического деления у такой зиготы с трисомией может приводить к таким кариотипам, как 94,XXXX,16+,16+.
Структурные хромосомные аномалии
Согласно классическим исследованиям, частота структурных хромосомных аберраций в материале выкидышей составляет 4—5%. Однако многие исследования были сделаны до широкого использования метода G-бэндинга. Современные исследования указывают на более высокую частоту структурных хромосомных аномалий у абортусов. Обнаруживаются самые разные виды структурных аномалий. Примерно в половине случаев эти аномалии являются унаследованными от родителей, примерно в половине случаев они возникают de novo.
Влияние хромосомных аномалий на развитие зиготы
Хромосомные аномалии зиготы проявляются как правило уже в первые недели развития. Выяснение конкретных проявлений каждой аномалии сопряжено с целым рядом трудностей.
Во многих случаях установление срока беременности при анализе материала выкидышей крайне затруднено. Обычно сроком зачатия считается 14-й день цикла, но у женщин с невынашиванием беременности часто бывают задержки цикла. Кроме того, очень трудно бывает установить дату «смерти» плодного яйца, поскольку от момента гибели до выкидыша может пройти много времени. В случыае триплоидии этот период может составить 10—15 недель. Применение гормональных препаратов может еще более удлиннить это время.
С учетом этих оговорок, можно сказать, что чем меньше срок беременности на момент гибели плодного яйца, тем выше частота хромосомных аберраций. Согласно исследованиям Кризи и Лоритсена, при выкидышах до 15 недель беременности частота хромосомных аберраций составляет около 50%, при сроке 18 — 21 неделя — около 15%, при сроке более 21 недели — около 5—8%, что примерно соответствует частоте хромосомных аберраций в исследованиях перинатальной смертности.
Фенотипические проявления некоторых летальных хромосомных аберраций
Моносомии X обычно останавливаются в развитии к 6 неделям после зачатия. В двух третях случаев плодный пузырь размером 5—8 см не содержит зародыша, но существует шнурообразное образование с элементами эмбриональной ткани, остатками желточного мешка, плацента содержит субамниотические тромбы. В одной трети случаев плацента имеет такие же изменения, но обнаруживается морфологически неизмененный зародыш, погибший в возрасте 40—45 дней после зачатия.
При тетраплоидиях развитие останавливается к сроку 2-3 недели после зачатия, морфологически эта аномалия характеризуется «пустым плодным мешком».
При трисомиях наблюдаются различные типы аномалий развития, в зависимости от того, какая хромосома является лишней. Однако в подавляющем большинстве случаев развитие останавливается на очень ранних сроках, элементов зародыша не обнаруживается. Это классический случай «пустого плодного яйца» (анэмбрионии).
Трисомия 16, очень частая аномалия, характеризуется наличием маленького плодного яйца диаметром около 2,5 см, в полости хориона находится небольшой амниотический пузырек около 5 мм в диаметре и эмбриональный зачаток размером 1—2 мм. Чаще всего развитие останавливается на стадии эмбрионального диска.
При некоторых трисомиях, например, при трисомиях 13 и 14, возможно развитие зародыша до срока около 6 недель. Зародыши характеризуются циклоцефалической формой головы с дефектами закрытия верхнечелюстных холмиков. Плаценты гипопластичны.
Зародыши с трисомиями 21 (синдром Дауна у новорожденных) не всегда имеют аномалии развития, а если и имеют, то незначительные, не могущие служить причиной их гибели. Плаценты в таких случаев бывают бедны клетками, и представляются остановившимися в развитии на ранней стадии. Гибель зародыша в таких случаях представляется следствием плацентарной недостаточности.
Заносы. Сравнительный анализ цитогенетических и морфологических данных позволяет выделить два типа заносов: классический пузырный занос и эмбриональный триплоидный занос.
Выкидыши при триплоидиях имеют четкую морфологическую картину. Это выражается в сочетании полной или (чаще) частичной пузырной дегенерации плаценты и амниотического пузырька с зародышем, размеры которого (зародыша) очень малы по сравнению с относительно большим амниотическим пузырьком. Гистологическое исследование показывает не гипертрофию, а гипотрофию пузырно измененного трофобласта, образующего микрокисты в результате многочисленный инвагинаций.
Напротив, классический пузырный занос не затрагивает ни амниотический мешок, ни зародыш. В пузырьках обнаруживается избыточное образование синцитиотрофобласта с выраженной васкуляризацией. Цитогенетически большинство классических пузырных заносов имеет кариотип 46,XX. Проведенные исследования позволили установить хромосомные сбои, участвующие в образовании пузырного заноса. Было показано, что 2 X-хромосомы в классическом пузырном заносе идентичны и имеют отцовское происхождение. Наиболее вероятным механизмом развития пузырного заноса является истинный андрогенез, возникающий вследствие оплодотворения яйцеклетки диплоидным сперматозоидом, возникшим в результате сбоя второго мейотического деления и последующим полным выключением хромосомного материала яйцеклетки. С точки зрения патогенеза, такие хромосомные нарушения близки к нарушениям при триплоидии.
Оценка частоты хромосомных нарушений в момент зачатия
Можно попробовать расчитать количество зигот с хромосомными аномалиями при зачатии, основываясь на частоте хромосомных аномалий, обнаруживаемых в материале выкидышей. Однако прежде всего следует отметить, что поразительное сходство результатов исследований материала выкидышей, проведенное в разных частях света, говорит о том, что хромосомные сбои в момент зачатия являются очень характерным явлением в репродукции у человека. Кроме того, можно констатировать, что реже всего встречающиеся аномалии (например, трисомии A, B и F) связаны с остановкой развития на очень ранних стадиях.
Анализ относительной частоты различных аномалий, возникающих при нерасхождении хромосом в процессе мейоза, позволяет сделать следующие важные выводы:
1. Единственной моносомией, обнаруживаемой в материале выкидышей, является моносомия X (15% всех аберраций). Напротив, аутосомные моносомии практически не обнаруживаются в материале выкидышей, хотя теоретически их должно быть столько же, сколько и аутосомных трисомий.
2. В группе аутосомных трисомий частота трисомий разных хромосом значительно варьирует. Исследования, выполненные с использованием метода G-бэндинга, позволили установить, что все хромосомы могут быть участницами трисомии, однако некоторые трисомии встречаются гораздо чаще, например, трисомия 16 встречается в 15% случаев всех трисомий.
Из этих наблюдений можно сделать вывод, что, скорее всего, частота нерасхождения разных хромосом приблизительно одинакова, а различная частота аномалий в материале выкидышей связана с тем, что отдельные хромосомные аберрации приводят к остановке развития на очень ранних стадиях и поэтому с трудом поддаются обнаружению.
Эти соображения позволяют приблизительно расчитать реальную частоту хромосомных нарушений в момент зачатия. Расчеты, сделанные Буэ, показали, что каждое второе зачатие дает зиготу с хромосомными аберрациями.
Данные цифры отражают среднюю частоту хромосомных аберраций при зачатии в популяции. Однако данные цифры могут значительно колебаться у разных супружеских пар. У некоторых супружеских пар вероятность возникновения хромосомных аберраций в момент зачатия значительно превышает средний риск в популяции. У таких супружеских пар невынашивание беременности на малых сроках происходит гораздо чаще, чем у остальных супружеских пар.
Данные расчеты подтверждаются другими исследованиями, проведенными с использованием других методов:
1. Классическими исследованиями Хертига
2. Определением уровня хорионического гормона (ХГ) в крови женщин после 10 после зачатия. Часто этот тест оказывается положительным, хотя менструация приходит вовремя или с небольшой задержкой, и субъективно наступления беременности женщина не замечает («биохимическая беременность»)
3. Хромосомный анализ материала, полученного при искусственных абортах показал, что при абортах на сроке 6—9 недель (4—7 недель после зачатия) частота хромосомных аберраций составляет примерно 8%, а при искусственных абортах на сроке 5 недель (3 недели после зачатия) эта частота возрастает до 25%.
4. Было показано, что нерасхождение хромосом в процессе сперматогенеза является очень частым явлением. Так Пирсон и сотр. обнаружили, что вероятность нерасхождения в процессе сперматогенеза для 1-й хромосомы составляет 3,5%, для 9-й хромосомы — 5%, для Y-хромосомы — 2%. Если и другие хромосомы имеют вероятность нерасхождения примерно такого же порядка, то тогда только 40% всех сперматозоидов имеют нормальный хромосомный набор.
Экспериментальные модели и сравнительная патология
Частота остановки развития
Хотя различия в типе плацентации и количестве плодов затрудняют сравнение риска неразвивающейся беременности у домашних животных и у человека, определенные аналогии проследить можно. У домашних животных процент летальных зачатий колеблется между 20 и 60%.
Изучение летальных мутаций у приматов дало цифры, сравнимые с таковыми у человека. Из 23 бластоцист, выделенных у макак до зачатия, у 10 были грубые морфологические аномалии.
Частота хромосомных аномалий
Только экспериментальные исследования позволяют провести хромосомный анализ зигот на разных стадиях развития и оценить частоту хромосомных аберраций. Классические исследования Форда выявили хромосомные аберрации у 2% зародышей мышей в возрасте от 8 до 11 дней после зачатия. Дальнейшие исследования показали, что это слишком продвинутая стадия развития зародышей, и что частота хромосомных аберраций гораздо выше (см. ниже).
Влияние хромосомных аберраций на развитие
Большой вклад в дело выяснения масштаба проблемы внесли исследования Альфреда Гроппа из Любека и Чарльза Форда из Оксфорда, проводившиеся на так называемых «табачных мышах» (Mus poschiavinus). Скрещивание подобных мышей с нормальными мышами дает большой спектр триплоидий и моносомий, позволяющих оценить влияние обоих типов аберраций на развитие.
Данные профессора Гроппа (1973 г.) приведены в таблице.
| Распределение эуплоидных и анэуплоидных зародышей у гибридных мышей | |||||
|---|---|---|---|---|---|
| Стадия развития | День | Кариотип | Всего | ||
| Моносомии | Эуплоидии | Трисомии | |||
| До имплантации | 4 | 55 | 74 | 45 | 174 |
| После имплантации | 7 | 3 | 81 | 44 | 128 |
| 9—15 | 3 | 239 | 94 | 336 | |
| 19 | 56 | 2 | 58 | ||
| Живые мыши | 58 | 58 | |||
Эти исследования позволили подтвердить гипотезу о равной вероятности возникновения моносомий и трисомий при зачатии: аутосомные моносомии возникают с такой же частотой, как и трисомии, но зиготы с аутосомными моносомиями погибают еще до имплантации и не обнаруживаются в материале выкидышей.
При трисомиях гибель зародышей происходит на более поздних сдадиях, но ни один зародыш при аутосомных трисомиях у мышей не доживает до родов.
Исследования группы Гроппа позволили показать, что в зависимости от типа трисомии, зародыши погибают на разных сроках: с трисомиями 8, 11, 15, 17 — до 12 дня после зачатия, с трисомиями 19 — ближе к сроку родов.
Патогенез остановки развития при хромосомных аномалиях
Исследование материала выкидышей показывает, что во многих случаях хромосомных аберраций эмбриогенез резко нарушается, так что элементов эмбриона не обнаруживается вообще («пустые плодные яйца», анэмбриония) (остановка развития до срока 2-3 недель после зачатия). В других случаях удается обнаружить элементы зародыша, часто неоформленные (остановка развития на сроке до 3-4 недель после зачатия). При наличии хромосомных аберраций эмбриогенез часто или вообще невозможен, или резко нарушается с самых ранних стадий развития. Проявления таких нарушений выражены в гораздо большей степени в случае аутосомных моносомий, когда развитие зиготы останавливается в первые дни после зачатия, но и в случае трисомий хромосом, имеющих ключевое значение для эмбриогенеза, развитие также прекращается в первые дни после зачатия. Так, например, трисомия 17 обнаруживается только у зигот, остановившихся в развитии на самых ранних стадиях. Кроме того, многие хромосомные аномалии связаны вообще с пониженной способностью к делению клеток, как показывает изучение культур таких клеток in vitro.
В других случаях развитие может продолжаться до 5—6—7 недель после зачатия, в редких случаях — дольше. Как показали исследования Филиппа, в таких случаях гибель плода объясняется не нарушением эмбрионального развития (обнаруживаемые дефекты сами по себе не могут быть причиной смерти зародыша), а нарушением формирования и функционирования плаценты (стадия развития плода опережает стадию формирования плаценты.
Исследования культур клеток плаценты при различных хромосомных аномалиях показали, что в большинстве случаев деление плацентарных клеток происходит гораздо медленнее, чем при нормальном кариотипе. Это во многом объясняет, почему новорожденные с хромосомными аномалиями обычно имеют низкую массу тела и сниженную массу плаценты.
Можно предположить, что многие нарушения развития при хромосомных аберрациях связаны именно с пониженной способностью клеток к делению. При этом возникает резкая диссинхронизация процессов развития зародыша, развития плаценты и индукции дифференциации и миграции клеток.
Недостаточное и запоздалое формирование плаценты может приводить к нарушению питания и к гипоксии зародыша, а также — к снижению гормональной продукции плаценты, что может быть дополнительной причиной развития выкидышей.
Исследования клеточных линий при трисомиях 13, 18 и 21 у новорожденных показало, что клетки делятся медленнее, чем при нормальном кариотипе, что проявляется в снижении плотности клеток в большинстве органов.
Загадкой является то, почему при единственной аутосомной трисомии, совместимой с жизнью (трисомия 21, синдром Дауна), в одних случаях происходит задержка развития зародыша на ранних стадиях и самопроизвольный выкидыш, а в других — ненарушенное развитие беременности и рождение жизнеспособного ребенка. Сравнение клеточных культур материала выкидышей и доношенных новорожденных при трисомии 21 показало, что различия в способности клеток к делению в первом и втором случаях резко различается, что возможно объясняет разную судьбу таких зигот.
Причины количественных хромосомных аберраций
Изучение причин хромосомных аберраций крайне затруднено, прежде всего из-за высокой частоты, можно сказать, всеобщности этого явления. Очень трудно корректно собрать контрольную группу беременных женщин, с большим трудом поддаются изучению нарушения сперматогенеза и оогенеза. Несмотря на это, некоторые этиологические факторы повышения риска хромосомных аберраций выяснить удалось.
Факторы, напрямую связанные с родителями
Влияние возраста матери на вероятность рождения ребенка с трисомией 21 наводит на мысль о возможном влиянии возраста матери на вероятность возникновения летальных хромосомных аберраций у зародыша. Приводимая ниже таблица показывает связь возраста матери с кариотипом материала выкидышей.
| Средний возраст матери при хромосомных аберрациях абортусов | ||
|---|---|---|
| Кариотип | Число наблюдений | Средний возраст |
| Нормальный | 509 | 27,5 |
| Моносомия X | 134 | 27,6 |
| Триплоидии | 167 | 27,4 |
| Тетраплоидия | 53 | 26,8 |
| Аутосомные трисомии | 448 | 31,3 |
| Трисомии D | 92 | 32,5 |
| Трисомии E | 157 | 29,6 |
| Трисомии G | 78 | 33,2 |
Как видно из таблицы, не было обнаружено связи между возрастом матери и самопроизвольными выкидышами, связанными с моносомией X, триплоидией или тетраплоидией. Повышение среднего возраста матери отмечено для аутосомных трисомий в целом, но по разным группам хромосом цифры были получены разные. Однако общее число наблюдений в группах недостаточно, чтобы уверенно судить о каких-либо закономерностях.
Возраст матери в большей степени связан с повышенным риском выкидышей с трисомиями акроцентрических хромосом группы D (13, 14, 15) и G (21, 22), что совпадает и со статистикой хромосомных аберраций при мертворождениях.
Для некоторых случаев трисомий (16, 21) было определено происхождение лишней хромосомы. Оказалось, что возраст матери связан с повышением риска трисомий только в случае материнского происхождения лишней хромосомы. Не было обнаружено связи возраста отца с повышением риска трисомий.
В свете исследований на животных высказываются предположения о возможной связи старения гамет и задержки оплодотворения на риск возникновения хромосомных аберраций. Под старением гамет понимают старение сперматозоидов в половых путях женщины, старение яйцеклетки либо в результате перезрелости внутри фолликула или в результате задержки выхода яйцеклетки из фолликула, либо в результате трубной перезрелости (запоздалого оплодотворения в трубе). Скорее всего, подобные законы действуют и у человека, но достоверных подтверждений этого пока не получено.
Факторы окружающей среды
Было показано, что вероятность хромосомных аберраций при зачатии повышается у женщин, подвергшихся действию ионизирующей радиации. Предполагается связь между риском хромосомных аберраций и действием других факторов, в частности — химических.
Заключение
1. Не каждую беременность удается сохранить на малых сроках. В большом проценте случаев выкидыши обусловлены хромосомными нарушениями у плода, и родить живого ребенка невозможно. Гормональное лечение может отсрочить момент выкидыша, но не может помочь зародышу выжить.
2. Повышенная нестабильность генома супругов является одним из причинных факторов бесплодия и невынашивания беременности. Выявить такие супружеские пары помогает цитогенетическое обследование с анализом на хромосомные аберрации. В некоторых случаях повышенной нестабильности генома специальная антимутагенная терапия может помочь повысить вероятность зачатия здорового ребенка. В других случаях рекомендуется донорская инсеминация или использование донорской яйцеклетки.
3. При невынашивании беременности, обусловленном хромосомными факторами, организм женщины может «запомнить» неблагоприятный иммунологический ответ на плодное яйцо (иммунологический импринтинг). В таких случаях возможно развитие реакции отторжения и на зародыши, зачатые после донорской инсеминации или с использованием донорской яйцеклетки. В таких случаях рекомендуется проведение специального иммунологического обследования.